

Spatiotemporal resolution of germinal center Tfh cell differentiation and divergence from central memory CD4+ T cell fate
- PMID: 37330549
- PMCID: PMC10276816
- DOI: 10.1038/s41467-023-39299-3
Spatiotemporal resolution of germinal center Tfh cell differentiation and divergence from central memory CD4+ T cell fate
Abstract
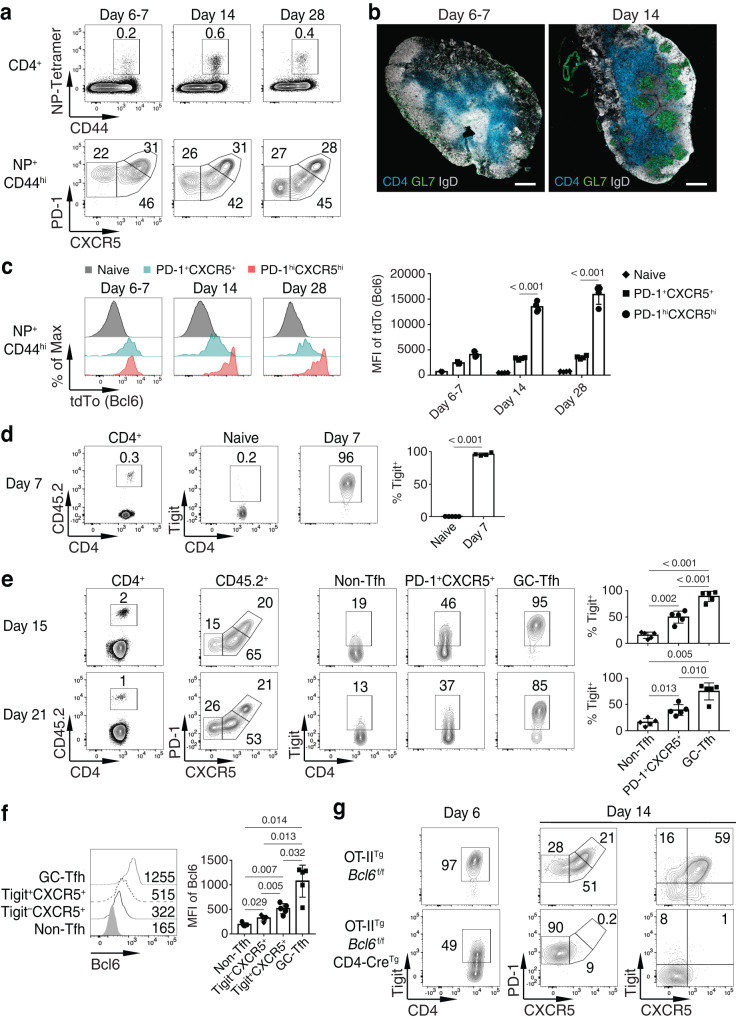
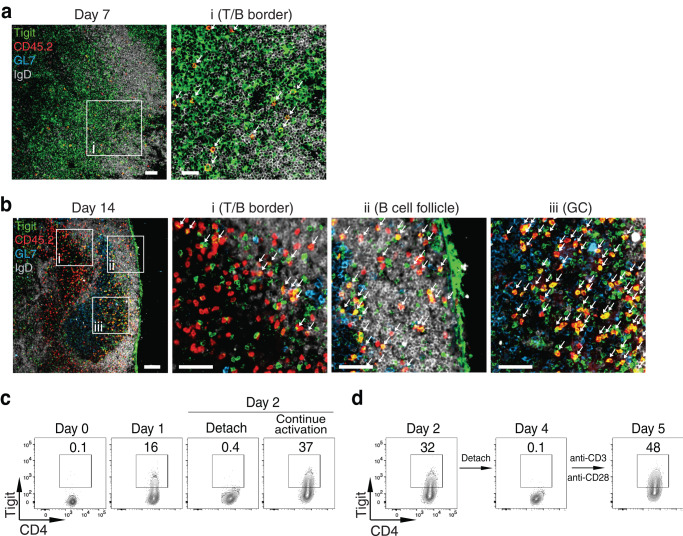
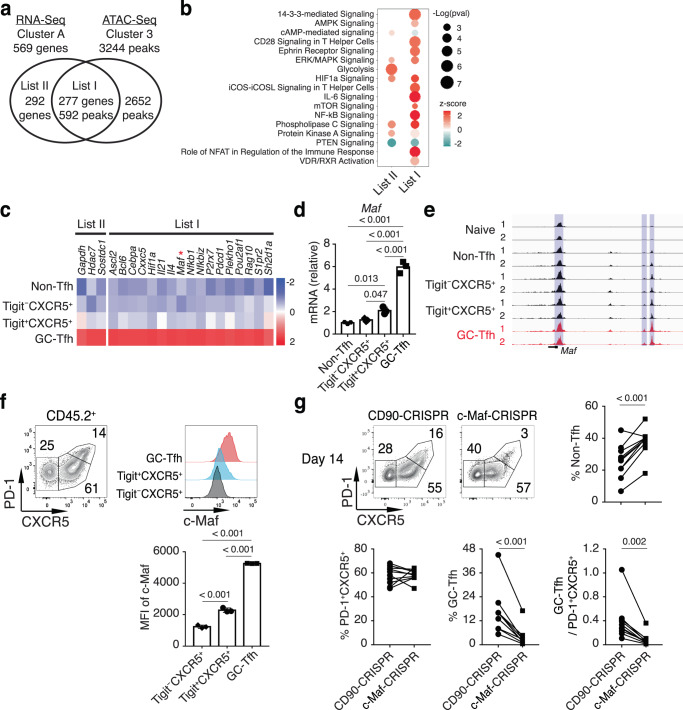
Follicular helper T (Tfh) cells are essential for germinal center (GC) B cell responses. However, it is not clear which PD-1+CXCR5+Bcl6+CD4+ T cells will differentiate into PD-1hiCXCR5hiBcl6hi GC-Tfh cells and how GC-Tfh cell differentiation is regulated. Here, we report that the sustained Tigit expression in PD-1+CXCR5+CD4+ T cells marks the precursor Tfh (pre-Tfh) to GC-Tfh transition, whereas Tigit-PD-1+CXCR5+CD4+ T cells upregulate IL-7Rα to become CXCR5+CD4+ T memory cells with or without CCR7. We demonstrate that pre-Tfh cells undergo substantial further differentiation at the transcriptome and chromatin accessibility levels to become GC-Tfh cells. The transcription factor c-Maf appears critical in governing the pre-Tfh to GC-Tfh transition, and we identify Plekho1 as a stage-specific downstream factor regulating the GC-Tfh competitive fitness. In summary, our work identifies an important marker and regulatory mechanism of PD-1+CXCR5+CD4+ T cells during their developmental choice between memory T cell fate and GC-Tfh cell differentiation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
- P30 AI027767/AI/NIAID NIH HHS/United States
- T32 AI007051/AI/NIAID NIH HHS/United States
- T32 AR069516/AR/NIAMS NIH HHS/United States
- R21 AI161774/AI/NIAID NIH HHS/United States
- R56 AI122842/AI/NIAID NIH HHS/United States
- R33 AI133679/AI/NIAID NIH HHS/United States
- R21 AI116188/AI/NIAID NIH HHS/United States
- P01 AI145815/AI/NIAID NIH HHS/United States
- T32 GM008361/GM/NIGMS NIH HHS/United States
- R61 AI133679/AI/NIAID NIH HHS/United States
- R56 AI167986/AI/NIAID NIH HHS/United States
- R01 AI130232/AI/NIAID NIH HHS/United States
- R33 AI116188/AI/NIAID NIH HHS/United States
- R01 AI122842/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
