

An in vitro workflow to create and modify infectious clones using replication cycle reaction
- PMID: 37331111
- PMCID: PMC10528026
- DOI: 10.1016/j.virol.2023.05.013
An in vitro workflow to create and modify infectious clones using replication cycle reaction
Abstract
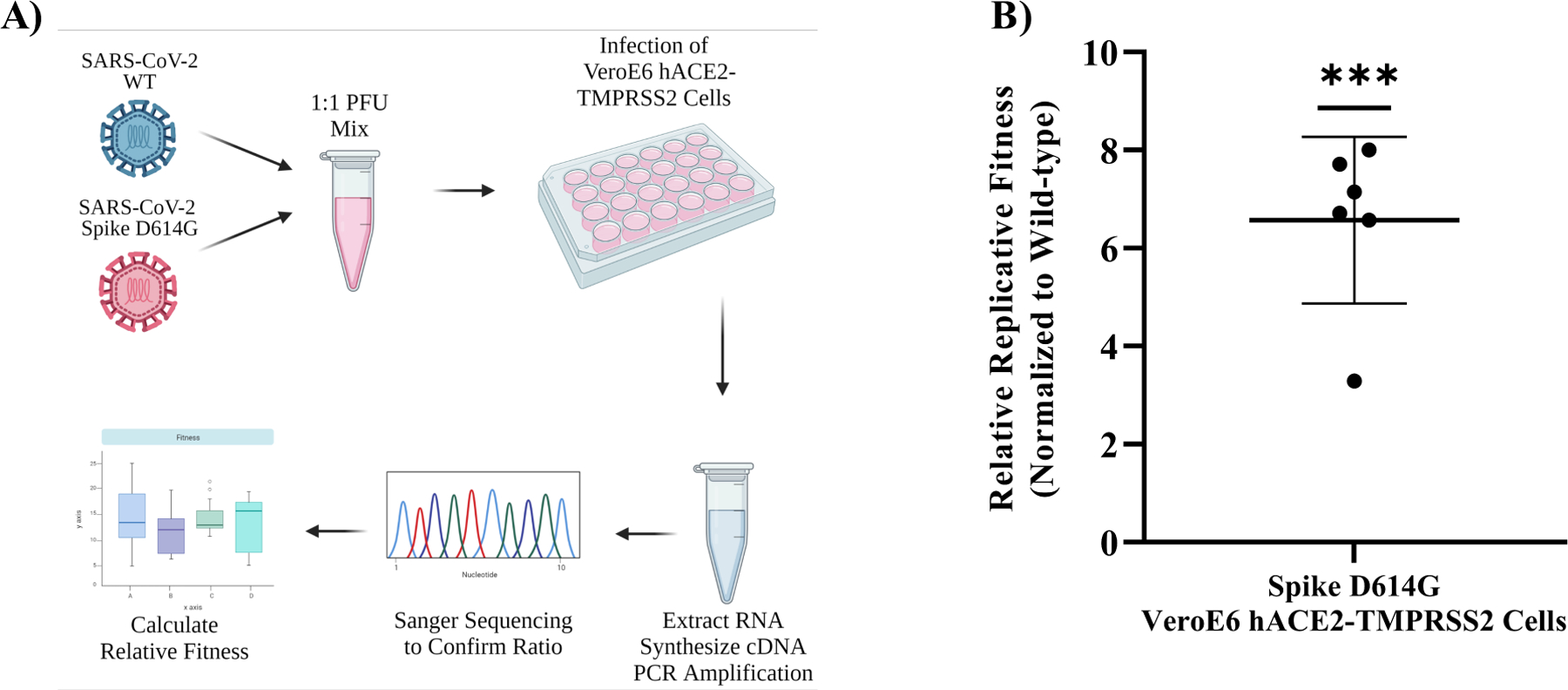
Reverse genetics systems are critical tools in combating emerging viruses which enable a better understanding of the genetic mechanisms by which viruses cause disease. Traditional cloning approaches using bacteria are fraught with difficulties due to the bacterial toxicity of many viral sequences, resulting in unwanted mutations within the viral genome. Here, we describe a novel in vitro workflow that leverages gene synthesis and replication cycle reaction to produce a supercoiled infectious clone plasmid that is easy to distribute and manipulate. We developed two infectious clones as proof of concept: a low passage dengue virus serotype 2 isolate (PUO-218) and the USA-WA1/2020 strain of SARS-CoV-2, which replicated similarly to their respective parental viruses. Furthermore, we generated a medically relevant mutant of SARS-CoV-2, Spike D614G. Results indicate that our workflow is a viable method to generate and manipulate infectious clones for viruses that are notoriously difficult for traditional bacterial-based cloning methods.
Keywords: Dengue virus; Infectious clones; Mutagenesis; Reverse genetics; SARS-CoV-2.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
