

This is a preprint.
Amyloid β Induces Lipid Droplet-Mediated Microglial Dysfunction in Alzheimer's Disease
- PMID: 37333071
- PMCID: PMC10274698
- DOI: 10.1101/2023.06.04.543525
Amyloid β Induces Lipid Droplet-Mediated Microglial Dysfunction in Alzheimer's Disease
Update in
-
Amyloid-β induces lipid droplet-mediated microglial dysfunction via the enzyme DGAT2 in Alzheimer's disease.Immunity. 2025 Jun 10;58(6):1536-1552.e8. doi: 10.1016/j.immuni.2025.04.029. Epub 2025 May 19. Immunity. 2025. PMID: 40393454
Abstract
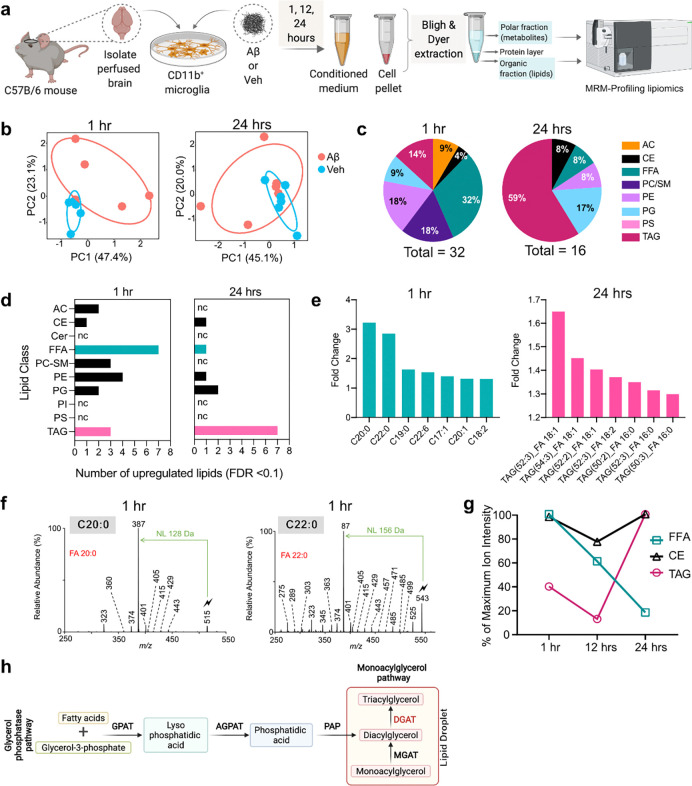
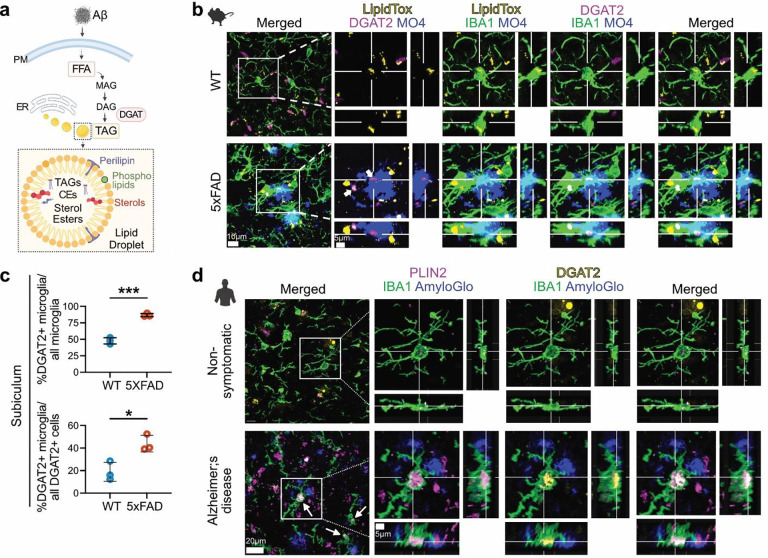
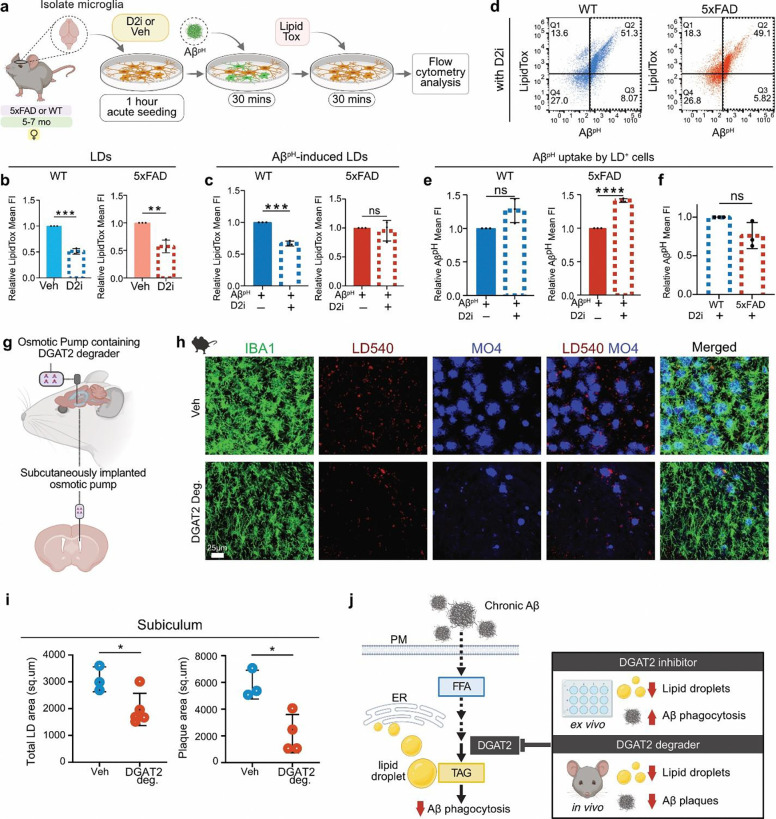
Several microglia-expressed genes have emerged as top risk variants for Alzheimer's disease (AD). Impaired microglial phagocytosis is one of the main proposed outcomes by which these AD-risk genes may contribute to neurodegeneration, but the mechanisms translating genetic association to cellular dysfunction remain unknown. Here we show that microglia form lipid droplets (LDs) upon exposure to amyloid-beta (Aβ), and that their LD load increases with proximity to amyloid plaques in brains from human patients and the AD mouse model 5xFAD. LD formation is dependent on age and disease progression and is prominent in the hippocampus in mice and humans. Despite differences in microglial LD load between brain regions and sexes in mice, LD-laden microglia exhibited a deficit in Aβ phagocytosis. Unbiased lipidomic analysis identified a decrease in free fatty acids (FFAs) and a parallel increase in triacylglycerols (TGs) as the key metabolic transition underlying LD formation. DGAT2, a key enzyme for converting FFAs to TGs, promotes microglial LD formation and is increased in 5xFAD and human AD brains. Inhibition or degradation of DGAT2 improved microglial uptake of Aβ and drastically reduced plaque load in 5xFAD mice, respectively. These findings identify a new lipid-mediated mechanism underlying microglial dysfunction that could become a novel therapeutic target for AD.
Keywords: Alzheimer’s disease; Microglia; glia; lipid droplets; lipidomics; lipids; metabolism; neurodegeneration; phagocytosis.
Conflict of interest statement
DECLARATION OF INTERESTS G.C. is the Director of the Merck-Purdue Center for Measurement Science funded by Merck Sharp & Dohme, a subsidiary of Merck and a co-founder of Meditati Inc., a startup developing smart drugs for mental health indications. The remaining authors declare no competing interests.
Figures


References
-
- Plascencia-Villa G. & Perry G. Status and future directions of clinical trials in Alzheimer’s disease. in International Review of Neurobiology vol. 154 3–50 (2020). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials