

This is a preprint.
13C metabolite tracing reveals glutamine and acetate as critical in vivo fuels for CD8+ T cells
- PMID: 37333111
- PMCID: PMC10274878
- DOI: 10.1101/2023.06.09.544407
13C metabolite tracing reveals glutamine and acetate as critical in vivo fuels for CD8+ T cells
Update in
-
13C metabolite tracing reveals glutamine and acetate as critical in vivo fuels for CD8 T cells.Sci Adv. 2024 May 31;10(22):eadj1431. doi: 10.1126/sciadv.adj1431. Epub 2024 May 29. Sci Adv. 2024. PMID: 38809979 Free PMC article.
Abstract
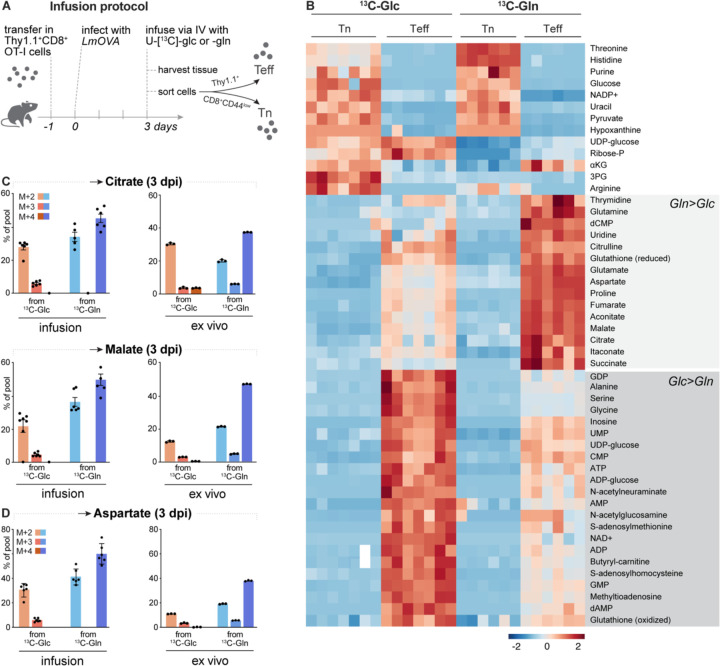
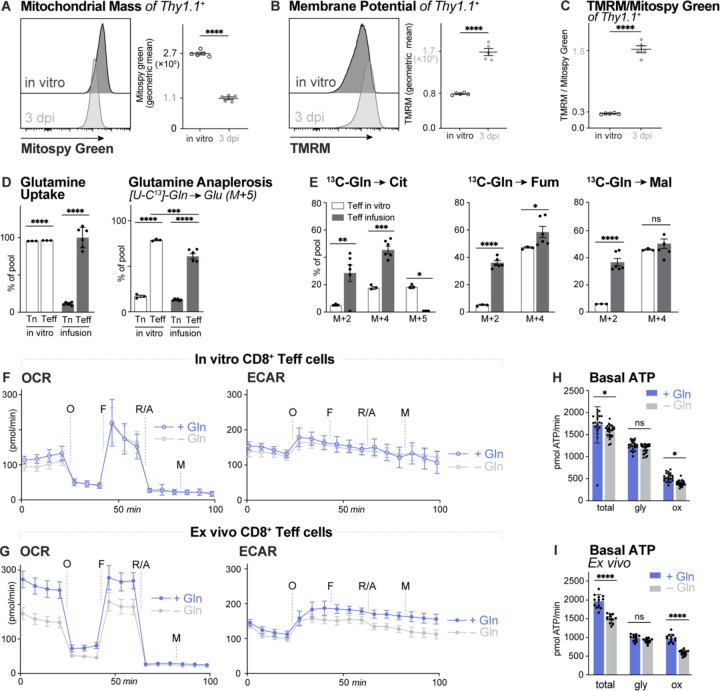
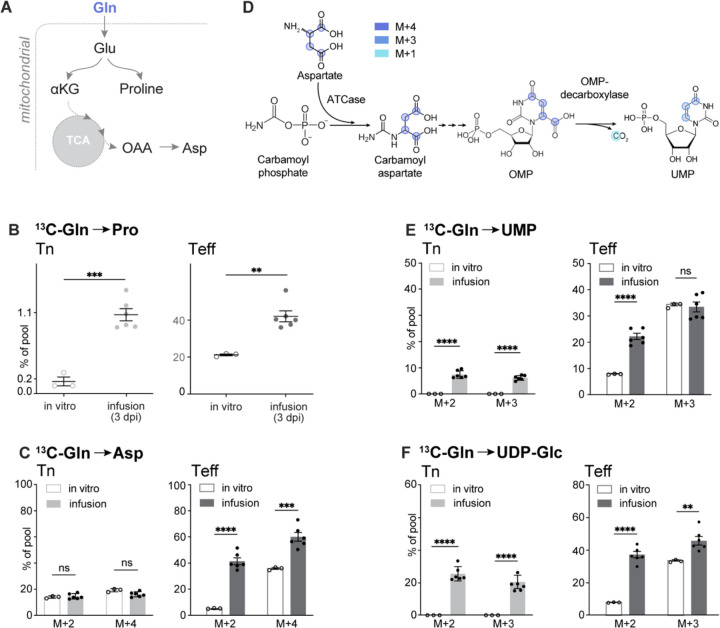
Infusion of 13C-labeled metabolites provides a gold-standard for understanding the metabolic processes used by T cells during immune responses in vivo. Through infusion of 13C-labeled metabolites (glucose, glutamine, acetate) in Listeria monocytogenes (Lm)-infected mice, we demonstrate that CD8+ T effector (Teff) cells utilize metabolites for specific pathways during specific phases of activation. Highly proliferative early Teff cells in vivo shunt glucose primarily towards nucleotide synthesis and leverage glutamine anaplerosis in the tricarboxylic acid (TCA) cycle to support ATP and de novo pyrimidine synthesis. Additionally, early Teff cells rely on glutamic-oxaloacetic transaminase 1 (Got1)-which regulates de novo aspartate synthesis-for effector cell expansion in vivo. Importantly, Teff cells change fuel preference over the course of infection, switching from glutamine- to acetate-dependent TCA cycle metabolism late in infection. This study provides insights into the dynamics of Teff metabolism, illuminating distinct pathways of fuel consumption associated with Teff cell function in vivo.
Conflict of interest statement
Competing interests: RGJ is a scientific advisor for Agios Pharmaceuticals and Servier Pharmaceuticals and is a member of the Scientific Advisory Board of Immunomet Therapeutics.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials