

LonP1 Links Mitochondria-ER Interaction to Regulate Heart Function
- PMID: 37333972
- PMCID: PMC10275618
- DOI: 10.34133/research.0175
LonP1 Links Mitochondria-ER Interaction to Regulate Heart Function
Abstract
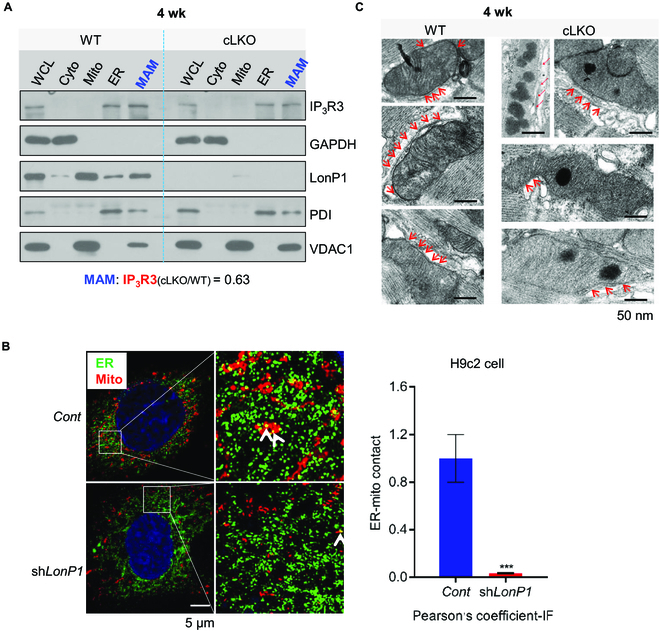
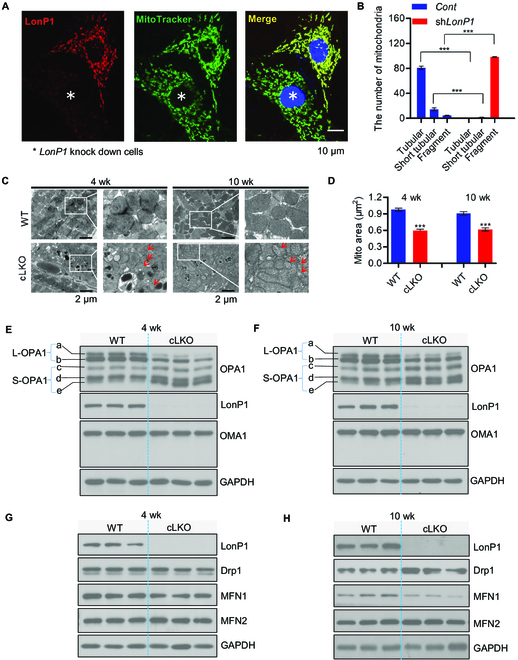
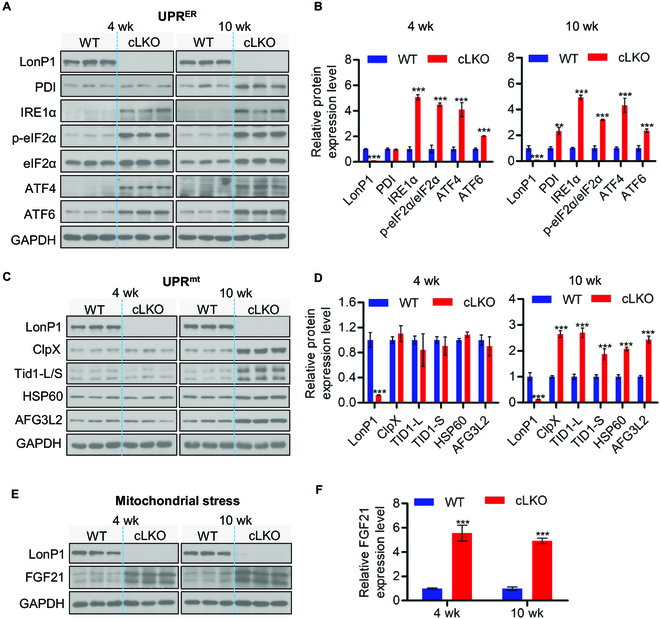
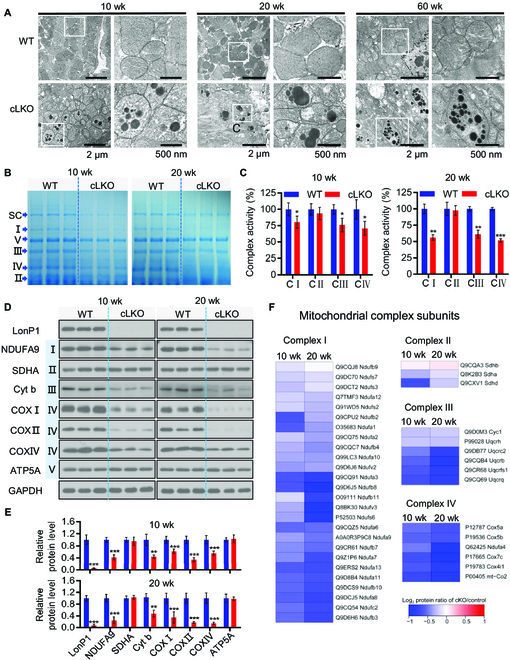
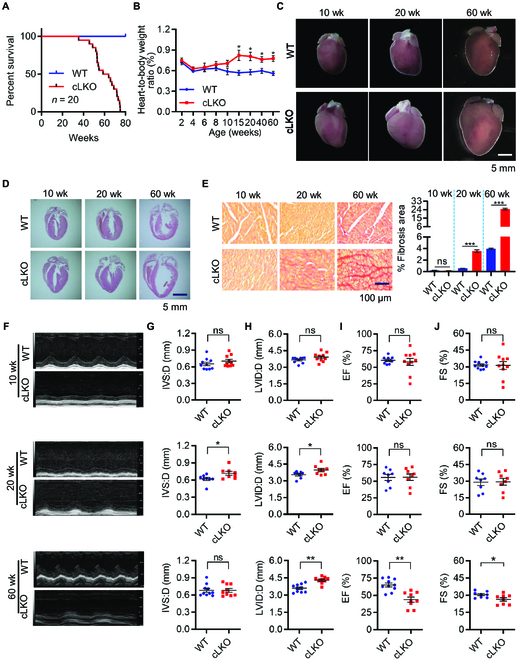
Interorganelle contacts and communications are increasingly recognized to play a vital role in cellular function and homeostasis. In particular, the mitochondria-endoplasmic reticulum (ER) membrane contact site (MAM) is known to regulate ion and lipid transfer, as well as signaling and organelle dynamics. However, the regulatory mechanisms of MAM formation and their function are still elusive. Here, we identify mitochondrial Lon protease (LonP1), a highly conserved mitochondrial matrix protease, as a new MAM tethering protein. The removal of LonP1 substantially reduces MAM formation and causes mitochondrial fragmentation. Furthermore, deletion of LonP1 in the cardiomyocytes of mouse heart impairs MAM integrity and mitochondrial fusion and activates the unfolded protein response within the ER (UPRER). Consequently, cardiac-specific LonP1 deficiency causes aberrant metabolic reprogramming and pathological heart remodeling. These findings demonstrate that LonP1 is a novel MAM-localized protein orchestrating MAM integrity, mitochondrial dynamics, and UPRER, offering exciting new insights into the potential therapeutic strategy for heart failure.
Copyright © 2023 Yujie Li et al.
Figures

References
-
- Pickett CB, Montisano D, Eisner D, Cascarano J. The physical association between rat liver mitochondria and rough endoplasmic reticulum. I. Isolation, electron microscopic examination and sedimentation equilibrium centrifugation analyses of rough endoplasmic reticulum-mitochondrial complexes. Exp Cell Res. 1980;128:343–352. - PubMed
LinkOut - more resources
Full Text Sources
