

Induced Pluripotent Stem Cell-Derived Extracellular Vesicles Promote Wound Repair in a Diabetic Mouse Model via an Anti-Inflammatory Immunomodulatory Mechanism
- PMID: 37335811
- PMCID: PMC10592465
- DOI: 10.1002/adhm.202300879
Induced Pluripotent Stem Cell-Derived Extracellular Vesicles Promote Wound Repair in a Diabetic Mouse Model via an Anti-Inflammatory Immunomodulatory Mechanism
Abstract
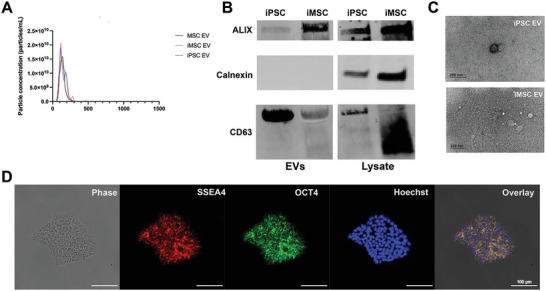
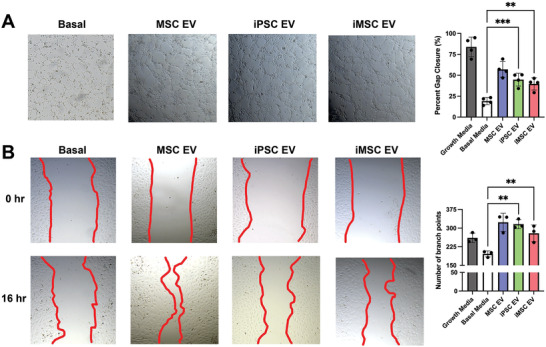
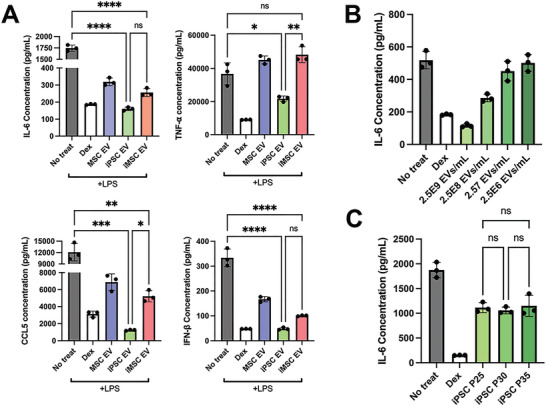
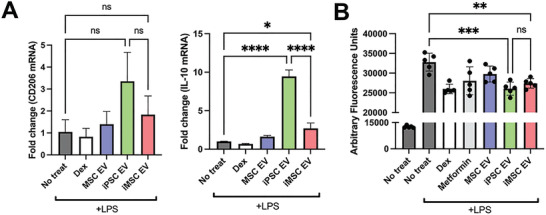
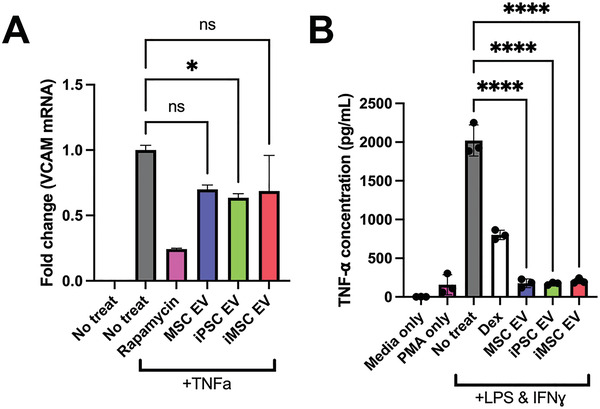
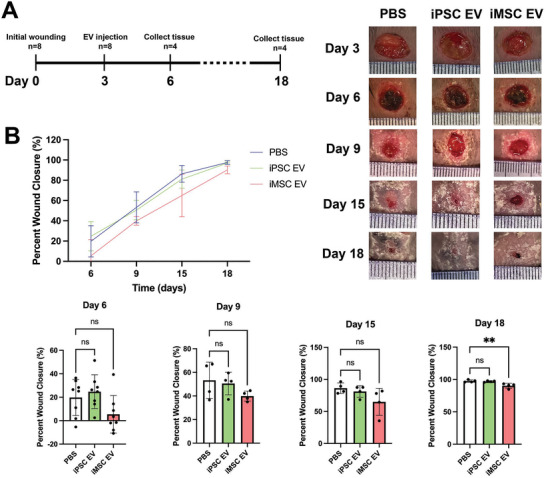
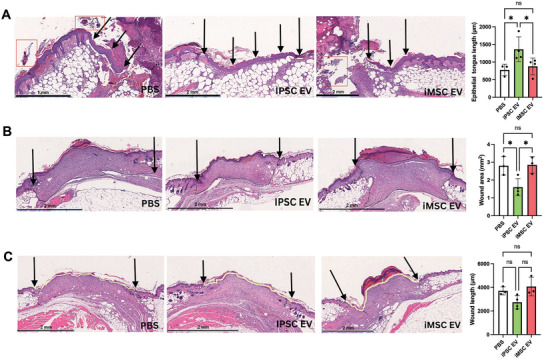
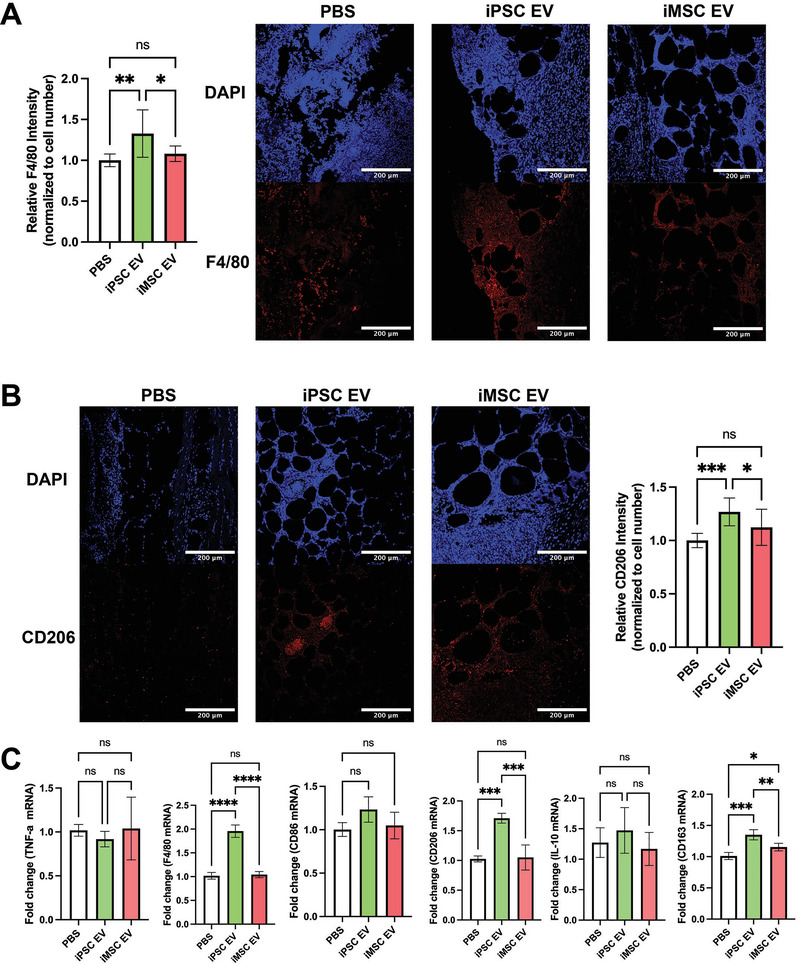
Extracellular vesicles (EVs) derived from mesenchymal stem/stromal cells (MSCs) have recently been explored in clinical trials for treatment of diseases with complex pathophysiologies. However, production of MSC EVs is currently hampered by donor-specific characteristics and limited ex vivo expansion capabilities before decreased potency, thus restricting their potential as a scalable and reproducible therapeutic. Induced pluripotent stem cells (iPSCs) represent a self-renewing source for obtaining differentiated iPSC-derived MSCs (iMSCs), circumventing both scalability and donor variability concerns for therapeutic EV production. Thus, it is initially sought to evaluate the therapeutic potential of iMSC EVs. Interestingly, while utilizing undifferentiated iPSC EVs as a control, it is found that their vascularization bioactivity is similar and their anti-inflammatory bioactivity is superior to donor-matched iMSC EVs in cell-based assays. To supplement this initial in vitro bioactivity screen, a diabetic wound healing mouse model where both the pro-vascularization and anti-inflammatory activity of these EVs would be beneficial is employed. In this in vivo model, iPSC EVs more effectively mediate inflammation resolution within the wound bed. Combined with the lack of additional differentiation steps required for iMSC generation, these results support the use of undifferentiated iPSCs as a source for therapeutic EV production with respect to both scalability and efficacy.
Keywords: exosomes; induced pluripotent stem cell-mesenchymal stem/stromal cells; induced pluripotent stem cells; inflammation; wound healing.
© 2023 The Authors. Advanced Healthcare Materials published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Update of
-
Induced pluripotent stem cell-derived extracellular vesicles promote wound repair in a diabetic mouse model via an anti-inflammatory immunomodulatory mechanism.bioRxiv [Preprint]. 2023 Mar 23:2023.03.19.533334. doi: 10.1101/2023.03.19.533334. bioRxiv. 2023. Update in: Adv Healthc Mater. 2023 Oct;12(26):e2300879. doi: 10.1002/adhm.202300879. PMID: 36993554 Free PMC article. Updated. Preprint.
References
-
- Nagelkerke A., Ojansivu M., Van Der Koog L., Whittaker T. E., Cunnane E. M., Silva A. M., Dekker N., Stevens M. M., Adv. Drug Delivery Rev. 2021, 175, 113775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
