

Reorganization of seagrass communities in a changing climate
- PMID: 37336970
- PMCID: PMC10356593
- DOI: 10.1038/s41477-023-01445-6
Reorganization of seagrass communities in a changing climate
Abstract
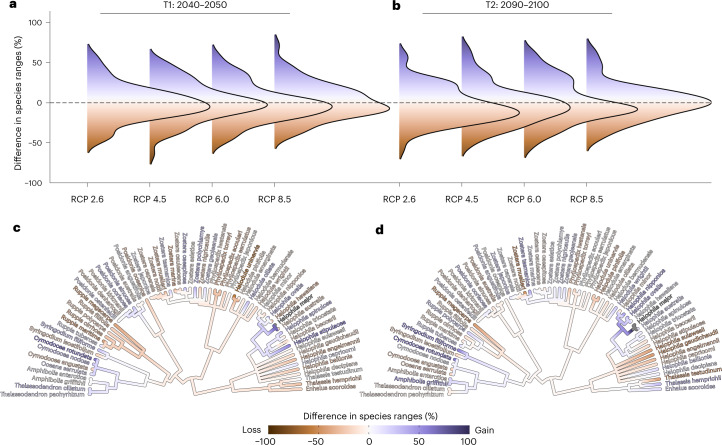
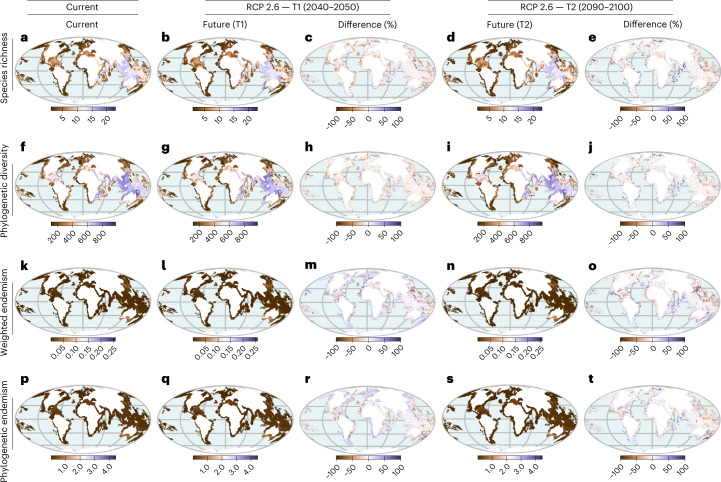
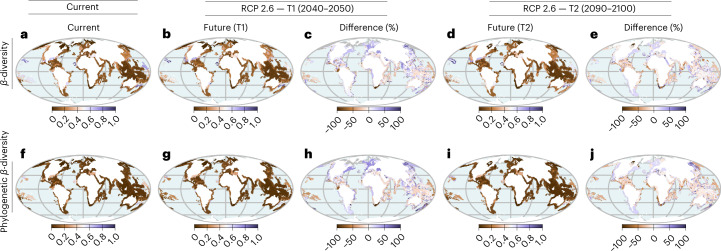
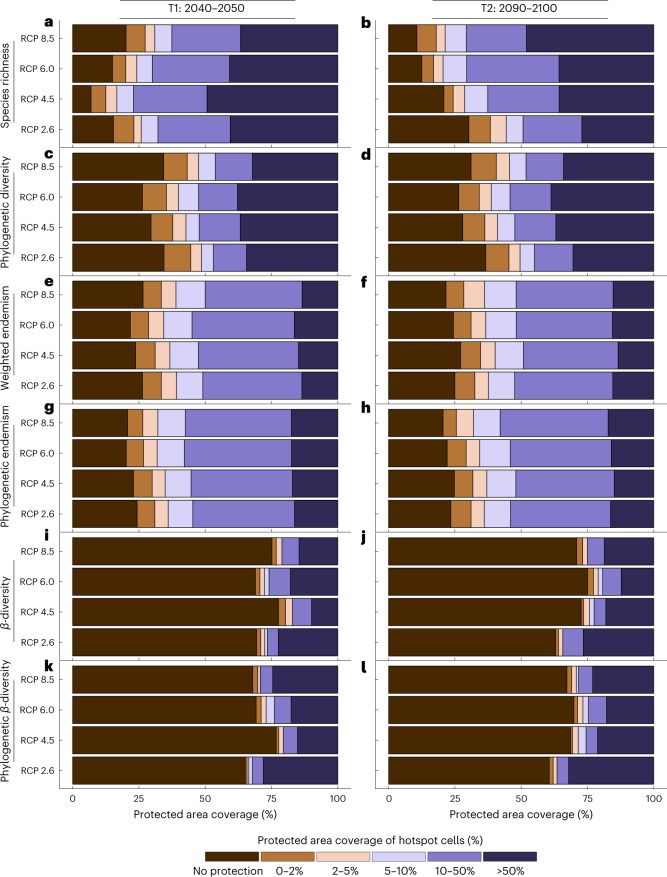
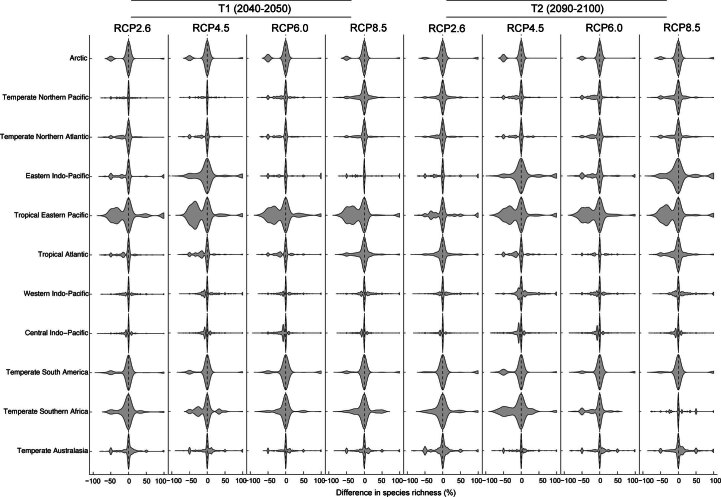
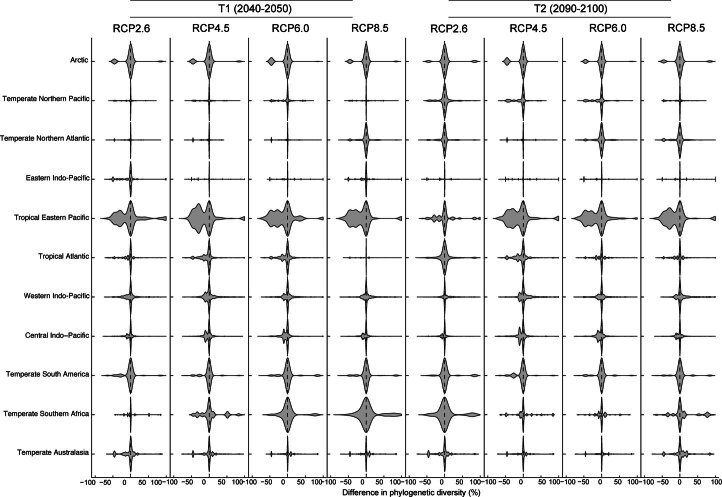
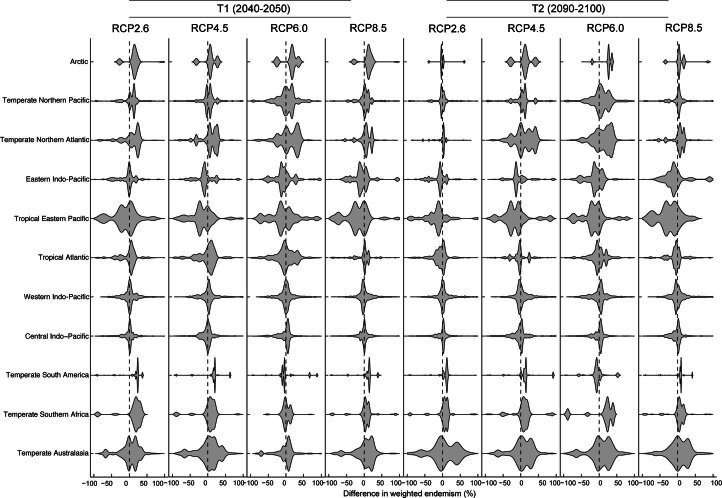
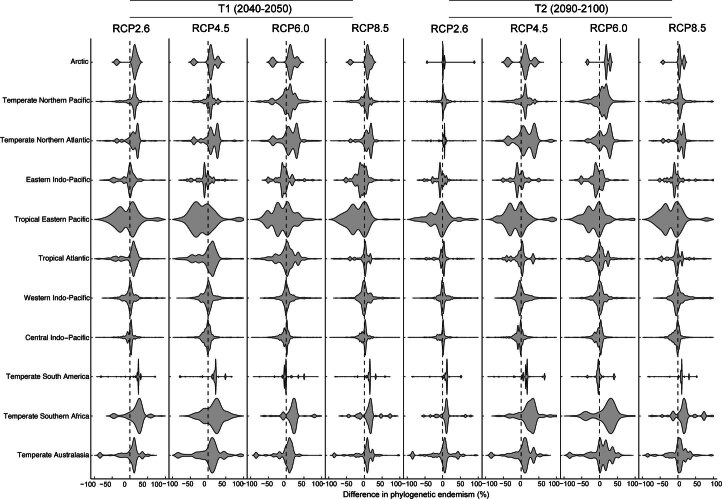
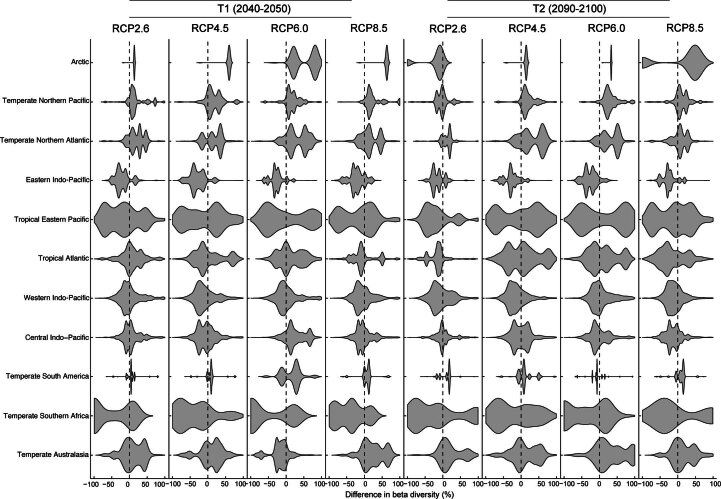
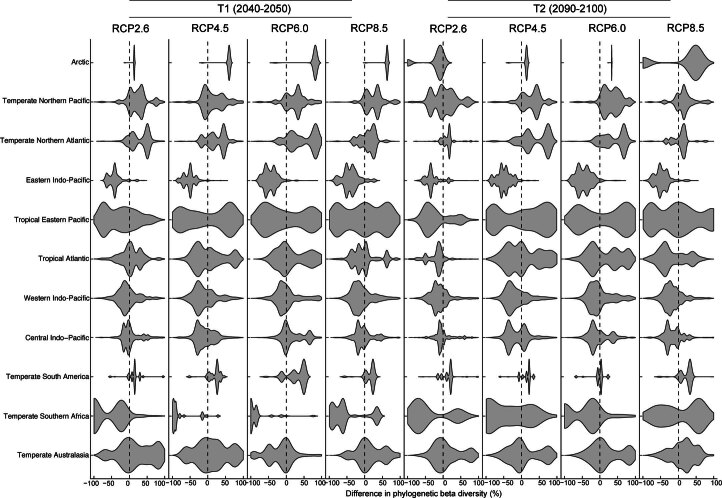
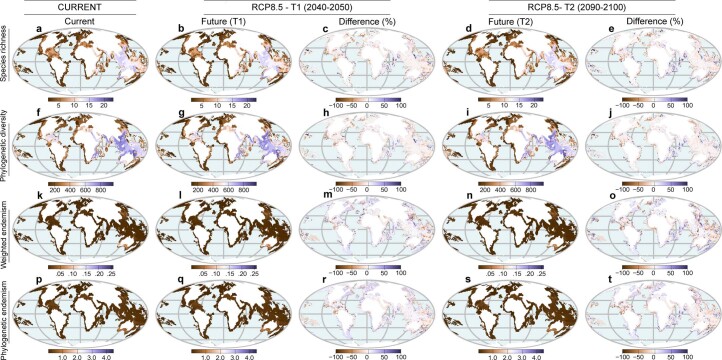
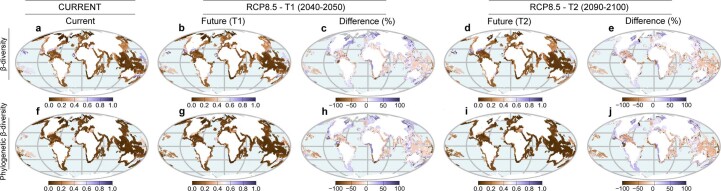
Although climate change projections indicate significant threats to terrestrial biodiversity, the effects are much more profound and striking in the marine environment. Here we explore how different facets of locally distinctive α- and β-diversity (changes in spatial composition) of seagrasses will respond to future climate change scenarios across the globe and compare their coverage with the existing network of marine protected areas. By using species distribution modelling and a dated phylogeny, we predict widespread reductions in species' range sizes that will result in increases in seagrass weighted and phylogenetic endemism. These projected increases of endemism will result in divergent shifts in the spatial composition of β-diversity leading to differentiation in some areas and the homogenization of seagrass communities in other regions. Regardless of the climate scenario, the potential hotspots of these projected shifts in seagrass α- and β-diversity are predicted to occur outside the current network of marine protected areas, providing new priority areas for future conservation planning that incorporate seagrasses. Our findings report responses of species to future climate for a group that is currently under represented in climate change assessments yet crucial in maintaining marine food chains and providing habitat for a wide range of marine biodiversity.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Misconception of model transferability precludes estimates of seagrass community reorganization in a changing climate.Nat Plants. 2024 Jul;10(7):1071-1074. doi: 10.1038/s41477-024-01735-7. Epub 2024 Jul 1. Nat Plants. 2024. PMID: 38951688 No abstract available.
References
-
- Burrows MT, et al. The pace of shifting climate in marine and terrestrial ecosystems. Science. 2011;334:652–655. - PubMed
-
- Blowes SA, et al. The geography of biodiversity change in marine and terrestrial assemblages. Science. 2019;366:339–345. - PubMed
-
- Jabbour J, Hunsberger C. Visualizing relationships between drivers of environmental change and pressures on land-based ecosystems. Nat. Resour. 2014;5:146–160.
-
- Elahi R, et al. Recent trends in local-scale marine biodiversity reflect community structure and human impacts. Curr. Biol. 2015;25:1938–1943. - PubMed
-
- Zeebe R, Ridgwell A, Zachos J. Anthropogenic carbon release rate unprecedented during the past 66 million years. Nat. Geosci. 2016;9:325–329.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
