

Enhancers of the PAIR4 regulatory module promote distal VH gene recombination at the Igh locus
- PMID: 37337907
- PMCID: PMC10390877
- DOI: 10.15252/embj.2022112741
Enhancers of the PAIR4 regulatory module promote distal VH gene recombination at the Igh locus
Abstract
While extended loop extrusion across the entire Igh locus controls VH -DJH recombination, local regulatory sequences, such as the PAIR elements, may also activate VH gene recombination in pro-B-cells. Here, we show that PAIR-associated VH 8 genes contain a conserved putative regulatory element (V8E) in their downstream sequences. To investigate the function of PAIR4 and its V8.7E, we deleted 890 kb containing all 14 PAIRs in the Igh 5' region, which reduced distal VH gene recombination over a 100-kb distance on either side of the deletion. Reconstitution by insertion of PAIR4-V8.7E strongly activated distal VH gene recombination. PAIR4 alone resulted in lower induction of recombination, indicating that PAIR4 and V8.7E function as one regulatory unit. The pro-B-cell-specific activity of PAIR4 depends on CTCF, as mutation of its CTCF-binding site led to sustained PAIR4 activity in pre-B and immature B-cells and to PAIR4 activation in T-cells. Notably, insertion of V8.8E was sufficient to activate VH gene recombination. Hence, enhancers of the PAIR4-V8.7E module and V8.8E element activate distal VH gene recombination and thus contribute to the diversification of the BCR repertoire in the context of loop extrusion.
Keywords: CTCF; Igh VH gene recombination; PAIR4 element; Pax5; novel recombination enhancers.
© 2023 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
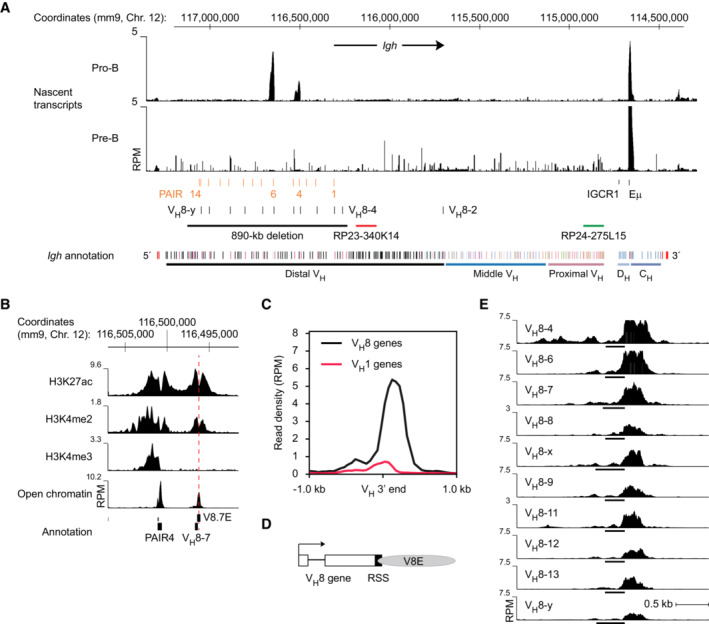
Mapping of total transcripts at the Igh locus in ex vivo sorted pro‐B and pre‐B cells. Total RNA was sequenced, as described in Materials and Methods, and plotted as reads per million mapped sequence reads (RPM). The locations of the PAIR elements, VH8 genes, IGCR1 sequence, and Eμ enhancer are shown together with the positions of the bacterial artificial chromosomes (BACs) RP23‐340K14 and RP24‐275L15 used as DNA‐FISH probes. The annotation of the C57BL/6 Igh locus indicates the distinct VH gene families (different colors) in the distal, middle, and proximal VH gene regions (Johnston et al, 2006), the DH (light blue) and CH (blue) elements, as well as the 3′RR enhancers (red) in the 3′ proximal Igh domain, together with the mm9 genomic coordinates of mouse chromosome 12. A black line indicates the extent of the 890‐kb deletion present in the Igh ∆890 allele.
Presence of active histone marks and open chromatin at PAIR4 and the V8.7E element located immediately downstream of the VH8‐7 gene in pro‐B cells. Ex vivo sorted pro‐B cells were used for determining open chromatin by ATAC‐seq and short‐term cultured pro‐B cells for ChIP‐seq analysis of the indicated histone marks with modification‐specific antibodies. The peak of open chromatin at the V8.7E element is indicated by a dashed line.
Specific presence of an open chromatin peak downstream of members of the VH8 gene family. The density of cumulative ATAC‐seq reads located downstream of members of the VH8 and VH1 gene families is shown. The 3′ end of the VH genes was used as a reference point for sequence alignment.
Schematic diagram of a VH8 gene with its RSS sequence and downstream V8E element. Gray shading indicates the extent of the ATAC‐seq peak present at the V8E element.
Identification of different V8E elements by their open chromatin peaks that are located downstream of individual VH8 genes (indicated by black lines). As the previously identified VH3609.8pg.160 and VH3609.14pg.181 genes (Johnston et al, 2006) were not mapped in the mouse mm9 or mm10 genomes, we refer to these PAIR6‐ and PAIR13‐associated genes as VH8‐x and VH8‐y, respectively. These genes have the following coordinates: VH8‐x gene –
chr12:116642631‐116643076 (mm9) or chr12:115404420‐115404865 (mm10); VH8‐y gene –chr12:117046639‐117047071 (mm9) or chr12:115808428‐115808860 (mm10).
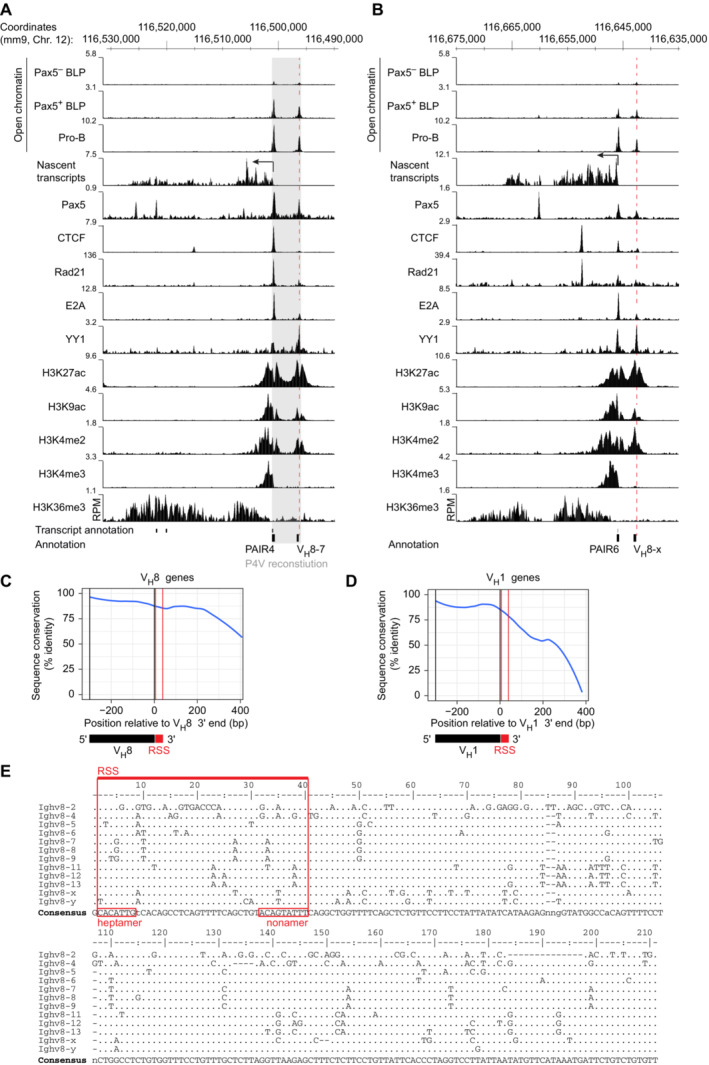
- A, B
Presence of active histone marks, transcription factor binding, open chromatin, and transcription in the PAIR4 (A) and PAIR6 (B) regions at the pro‐B cell stage. Open chromatin was furthermore mapped by ATAC‐seq in hCD2− (Pax5−) and hCD2+ (Pax5+) BLPs that were sorted from the bone marrow of Pax5 ihCd2/ihCd2 mice (Fuxa & Busslinger, ; Hill et al, 2020). The RPM scales for displaying the open chromatin peaks in BLPs and pro‐B cells were adjusted to show equal densities of the open chromatin peaks present at a gene‐dense genomic region, including the ubiquitously expressed Tbp locus. The location of PAIR4, PAIR6, and their associated VH8 genes are shown together with the exons of the PAIR4‐derived lncRNA (RIKEN clone CJ056205; below) and the mm9 genomic coordinates of mouse chromosome 12 (above). As the previously identified VH3609.8pg.160 (Johnston et al, 2006) was not mapped in the mouse mm9 or mm10 genome, we refer to this PAIR6‐associated gene as VH8‐x. The coordinates for the VH8‐x gene are
chr12:116642631‐116643076 (mm9) or chr12:115404420–115,404,865 (mm10). Gray overlay indicates the DNA sequences of the PAIR4‐V8.7E module that were inserted with the Floxin method at the deletion point of the Igh ∆890 allele to generate the Igh P4V allele (Fig 3A). RPM, reads per million mapped sequence reads. - C, D
Sequence conservation of the aligned VH8 (C) and VH1 (D) gene regions and their 3′ flanking sequences (~400 bp). The line denotes the LOESS‐smoothed position‐wise maximum sequence identity across the multiple sequence alignments. The coding sequences of the VH genes are indicated together with the recombination signal sequences (RSS).
- E
Sequence alignment of the ~200‐bp conserved region downstream of the 3′ end of the VH8 genes. Dots indicate identical nucleotides relative to the consensus sequence shown below. Nonidentical nucleotides are shown by their respective letters (C, G, T, and A). Gaps in the alignment are indicated by dashes. Numbers refer to the positions downstream of the VH8 3′ end.
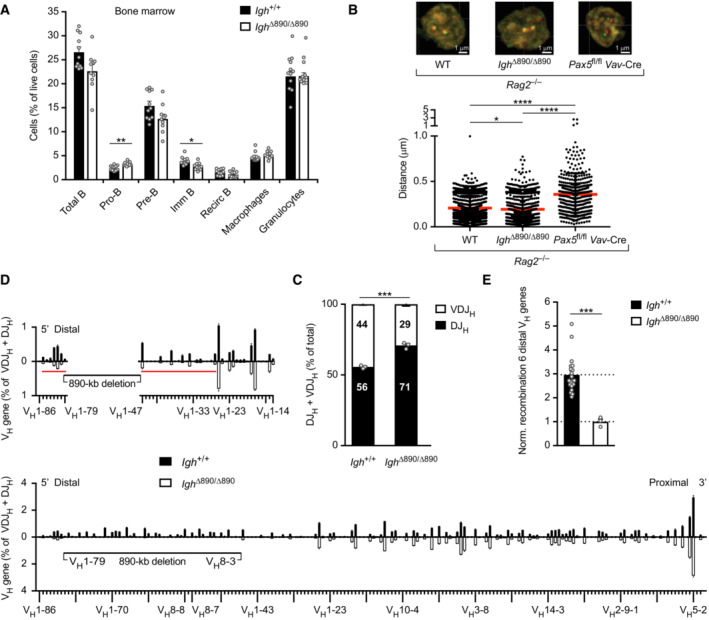
- A
B cell development in Igh ∆890/∆890 mice. The relative frequencies of the indicated cell types were determined by flow cytometric analysis of bone marrow cells from Igh ∆890/∆890 and Igh +/+ mice at the age of 3–4 weeks, indicating a 1.3‐fold increase of pro‐B cells and a moderate decrease at subsequent B cell developmental stages in Igh ∆890/∆890 mice. Statistical data are shown as a mean value with SEM.
- B
Two‐color 3D DNA‐FISH analysis of ex vivo sorted pro‐B cells of the indicated genotypes with the RP23‐340K14 (red) and RP24‐275L15 (green) BAC probes (Fig 1A). Representative images are shown above. Dot plots (below) show the distances measured between the two DNA signals in individual Igh alleles (2,123 for Rag2 −/−, 1,514 for Igh ∆890/∆890 Rag2 −/−, and 502 for Vav‐Cre Pax5 fl/fl (Pax5 ∆/∆) Rag2 −/− pro‐B cells) together with the mean distance (red bar) and SEM determined for each genotype. In total, four independent DNA‐FISH experiments were performed.
- C, D
VH gene recombination analysis in ex vivo sorted Igh ∆890/∆890 and Igh +/+ pro‐B cells, as determined by VDJ‐seq experiments (Chovanec et al, 2018). (C) The percentages of uniquely identified DJH and VDJH sequences are shown as mean percentages with SEM. Each dot corresponds to an independent VDJ‐seq experiment performed with sorted pro‐B cells from one mouse. (D) VDJ‐seq analysis of VH gene rearrangements in Igh +/+ pro‐B cells (black, above line) and Igh ∆890/∆890 pro‐B cells (white bars, below line). The different VH genes (horizontal axis) are aligned according to their position in the Igh locus (Dataset EV1). The usage of each VH gene (vertical axis) is shown as a percentage of all VDJH and DJH recombination events determined for each pro‐B cell type. The relative frequency of each VH gene is shown as a mean value with SEM and is based on three independent VDJ‐seq experiments for pro‐B cells of each genotype. The enlargement (upper part) highlights the differences in recombination frequency of the VH genes adjacent to the 890‐kb deletion. The red horizontal line indicates the extent of reduced VH gene recombination on either side of the deletion point in Igh ∆890/∆890 pro‐B cells.
- E
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) in Igh ∆890/∆890 and Igh +/+ pro‐B cells. The average recombination frequency of the six distal VH genes was calculated as mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1. The average recombination frequency in Igh +/+ pro‐B cells was determined based on all VDJ‐seq experiments that were performed in this study with this control pro‐B cell type. The average recombination frequency of each distal VH gene in Igh ∆890/∆890 and Igh +/+ pro‐B cells is shown in Fig EV3C.
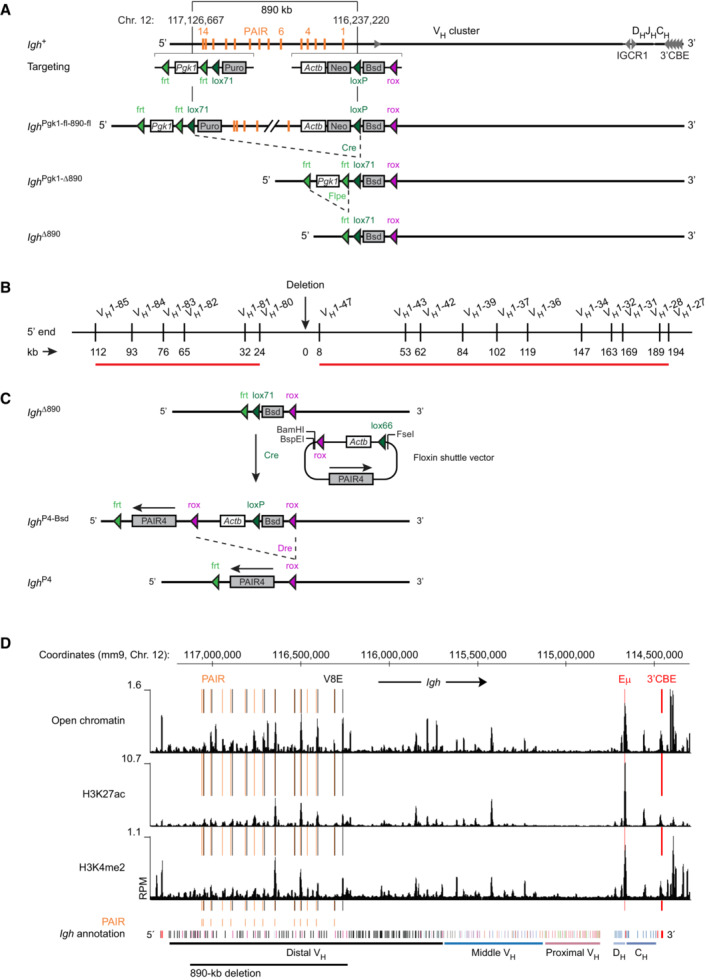
- A
Generation of the Igh ∆890 allele. The indicated selection cassettes were used for introducing the upstream lox71 site and the downstream loxP site at the specified positions of the Igh locus by sequential ES cell targeting to generate the Igh Pgk1‐fl‐890‐fl allele. The Igh ∆890 allele was subsequently generated by sequential deletion of the indicated sequences by Cre‐ and Flpe‐mediated recombination in vivo in the mouse, as described in Materials and Methods. Actb, human β‐actin promoter; Bsd, blasticidin resistance gene; Neo, neomycin resistance gene; Puro, puromycin resistance gene; Pgk1, phosphoglycerate kinase‐1 promoter.
- B
Location of the VH1 gene family members, which undergo rearrangements in Igh +/+ pro‐B cells and exhibit strongly reduced recombination (indicated by the red line; see Fig 2D) upon deletion of the 890‐kb region in the 5′ region of the Igh locus. The distance of the different VH genes from the deletion point is shown in kilobases (kb).
- C
Schematic diagram of the Floxin system used for inserting the different PAIR4 constructs and the Eμ enhancer at the deletion point of the Igh ∆890 allele in ES cells. See Materials and Methods for detailed description of the reconstitution experiments.
- D
Presence of active chromatin across the Igh locus in Rag2 −/− pro‐B cells. Open chromatin was mapped by ATAC‐seq, while the presence of H3K27ac and H3K4me2 peaks was identified by ChIP‐seq analysis with anti‐H3K27ac and anti‐H3K4me2 antibodies. The peaks corresponding to PAIR and V8E elements are highlighted by orange or black lines, respectively. The Eμ enhancer and 3′CBE region are indicated in red. The annotation of the C57BL/6 Igh locus with the different VH gene families (different colors), the DH (light blue), and CH (blue) elements is shown together with the extent of the 890‐kb deletion (black line) and the mm9 genomic coordinates of mouse chromosome 12.
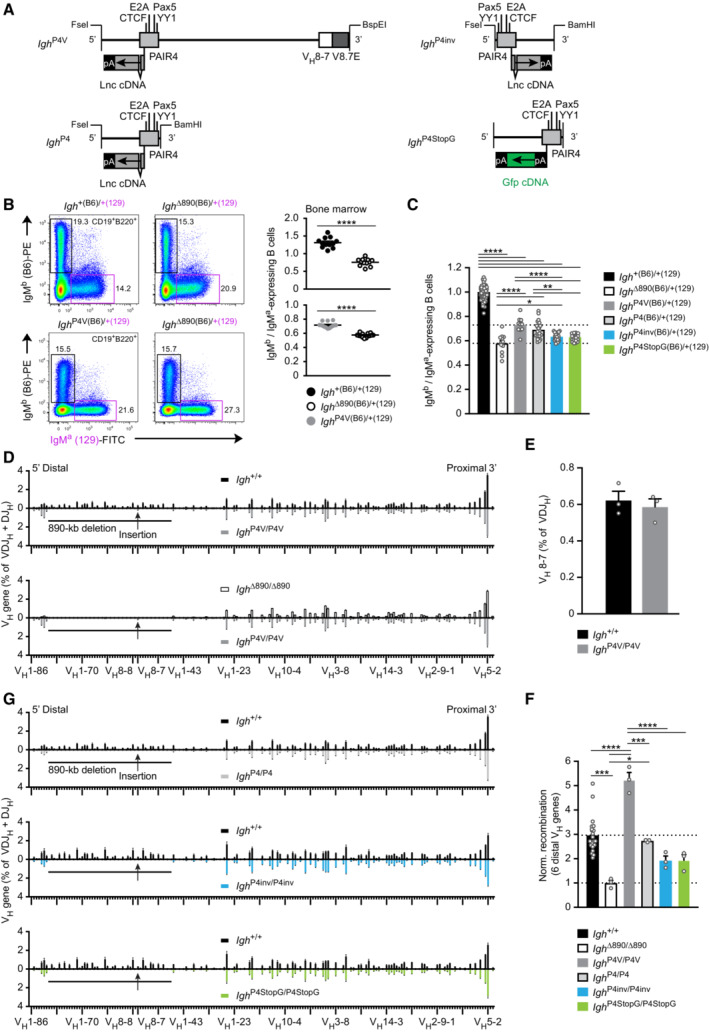
Schematic diagram of the different PAIR4 constructs used for reconstitution experiments. The PAIR4 element with its transcription factor binding sites and the cDNA insertion of the PAIR‐derived lncRNA (RIKEN clone CJ056205) in exon 2 of PAIR4 is shown together with the VH8‐7 gene and its downstream V8.7E element. Six copies of a synthetic polyadenylation sequence (pA; Levitt et al, 1989) linked to Gfp cDNA were inserted in exon 1 of PAIR4 to generate the P4StopG insert. FseI‐BspEI or FseI‐BamHI DNA fragments of the different PAIR4 constructs were inserted at the deletion point into the Igh ∆890 allele with the Floxin system (Singla et al, 2010) as described in detail in Fig EV2C and Materials and Methods. pA, 6 polyadenylation sites.
Flow cytometric determination of the ratio of immature IgMb (B6) to IgMa (129) B cells in the bone marrow of Igh ∆890(B6)/+(129), Igh P4V (B6)/+(129) and control Igh +(B6)/+(129) mice, which were generated by crossing Igh ∆890/+, Igh P4V/+ and Igh +/+ mice on the C57BL/6 (B6) background with Igh +/+ mice of the 129/Sv (129) strain. The rearranged Igh alleles of the C57BL/6 and 129/Sv strains give rise to the expression of IgMb and IgMa, respectively. The IgMb (B6) to IgMa (129) ratio is shown as a mean value with SEM (right).
Summary of the ratios of immature IgMb (B6) to IgMa (129) B cells, determined for the indicated six mouse strains analyzed and shown as mean values with SEM. The ratio determined for control Igh +(B6)/+(129) B cells was set to 1.
Comparison of the VDJ‐seq data obtained with pro‐B cells of the Igh +/+ (black), Igh ∆890/∆890 (white bars), and Igh P4V/P4V (gray) genotypes. The different VH genes (horizontal axis) are aligned according to their position in the Igh locus (Dataset EV1). The usage of each VH gene (vertical axis) is shown as a percentage of all VDJH and DJH recombination events determined for each pro‐B cell type. The relative frequency of each VH gene is shown as a mean value with SEM and is based on three independent VDJ‐seq experiments for pro‐B cells of each genotype.
Recombination frequency of the inserted VH8‐7 gene, which was determined as a percentage of all VDJH recombination events in Igh +/+ and Igh P4V/P4V pro‐B cells and is shown as a mean value with SEM.
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) determined in pro‐B cells of the indicated genotypes, based on the data shown in (D, G). The average recombination frequency of the six distal VH genes was calculated as a mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1. The average recombination frequency of each distal VH gene in pro‐B cells of the indicated genotypes is shown in Fig EV3C.
Comparison of the VDJ‐seq data obtained with pro‐B cells of the Igh +/+ (black), Igh P4/P4 (light gray), Igh P4inv/P4inv (blue), and Igh P4StopG/P4StopG (green) genotypes. For further description see (D).
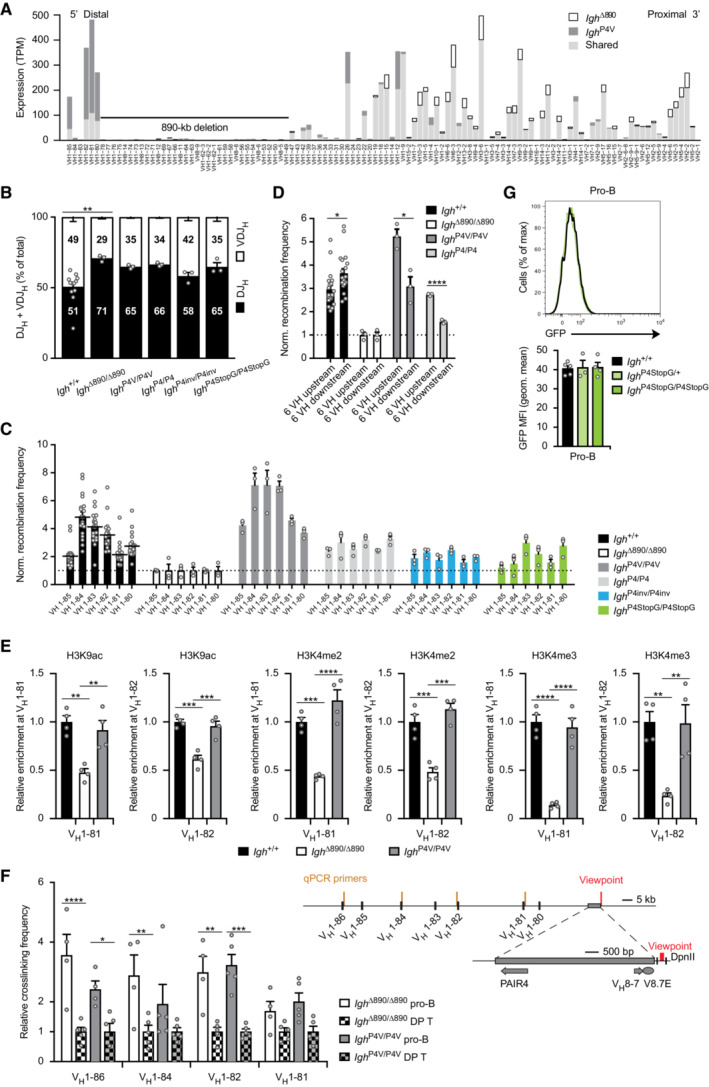
VH gene expression from the Igh P4V(B6) and Igh ∆890(B6) alleles in immature IgMb (B6) B cells sorted from the bone marrow of Igh P4V(B6)/+(129) and Igh ∆890(B6)/+(129) mice, respectively (see also Fig 3B). The expression (TPM) value of each VH gene is shown as shared (light gray) or unique expression of the Igh P4V(B6) allele (dark gray) or Igh ∆890(B6) allele (white bars). The expression data are shown as mean TPM values and are based on three or two independent RNA‐seq experiments for the immature B cells of Igh P4V(B6)/+(129) and Igh ∆890(B6)/+(129) mice, respectively. The different VH genes (horizontal axis) are aligned according to their position in the Igh locus (Dataset EV1). The 890‐kb deletion is indicated by a black line. TPM, transcripts per million; B6, C57BL/6 strain; 129, 129/Sv strain.
VDJ‐seq analysis of Igh rearrangements in pro‐B cells of the indicated genotypes. The percentages of uniquely identified DJH and VDJH sequences are shown as mean percentage with SEM. The high variance caused by the many data points obtained with control Igh +/+ pro‐B cells is likely responsible for the fact that only the comparison between Igh +/+ and Igh ∆890/∆890 pro‐B cells reached statistical significance.
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) determined in pro‐B cells of the indicated genotypes, based on the data shown in Fig 3D and G. The recombination frequency of each of the six distal VH genes was calculated as a mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1 (dashed line).
The PAIR4‐V8.7E module and PAIR4 elements differentially activate the recombination of VH genes located upstream or downstream of their insertion. The normalized recombination frequency of the first six upstream VH genes (VH1‐85 to VH1‐80) and the first six downstream VH genes (VH1‐47, VH1‐43, VH1‐42, VH1‐39, VH1‐37, VH1‐36; Fig EV2B) was determined in pro‐B cells of the indicated genotypes, based on the data shown in Fig 3D and G. The average recombination frequency of the six upstream and six downstream VH genes was calculated as mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1.
Active chromatin at the VH1‐81 and VH1‐82 genes in Igh P4V/P4V, Igh ∆890/∆890 and Igh +/+ pro‐B cells. Short‐term cultured pro‐B cells of the indicated genotypes were used for native ChIP analysis with anti‐H3K9ac, anti‐H3K4me2 and anti‐H3K4me3 antibodies. Input and precipitated DNA were quantified by qPCR analysis with primers amplifying the sequences of the VH1‐81, VH1‐82 and Tbp genes. The amount of precipitated DNA was calculated as mean percentage of input with SEM and was normalized to the value obtained for the positive Tbp control. The relative enrichment is shown by setting the normalized value of the control Igh +/+ pro‐B cells to 1.
3C‐qPCR analysis of the long‐range interactions between the PAIR4‐V8.7E module and distal VH genes in sorted bone marrow pro‐B cells and CD4+CD8+ double‐positive (DP) thymocytes from Igh P4V/P4V and Igh ∆890/∆890 mice. The relative crosslinking frequencies between the reference DpnII fragment (viewpoint, located downstream of the insertion site) and the indicated distal VH genes were determined with independently prepared 3C‐templates and were normalized relative to the crosslinking frequency measured for the control Ercc3 gene (see Materials and Methods). The mean crosslinking frequency with SEM is shown for each VH gene by setting the mean value obtained with DP T cells for each genotype to 1.
Absence of GFP expression in pro‐B cells of Igh P4StopG/+ (light green) and Igh P4StopG/P4StopG (green) mice compared with control Igh +/+ (black) mice, as indicated by flow‐cytometric analysis and geometric mean fluorescence intensity measurement. Geometric mean fluorescence intensity measurements are shown as a mean value with SEM.
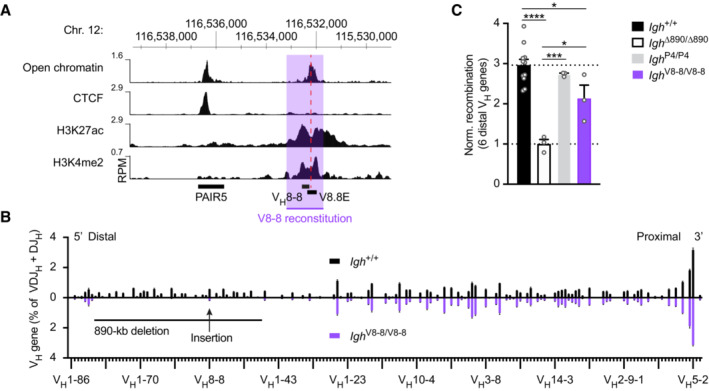
Presence of open chromatin and the active histone marks H3K27ac and H3K4me2 at the V8.8E enhancer in pro‐B cells. The location of the VH8‐8 gene, V8.8E enhancer (dashed line), and PAIR5 element are shown together with the mm9 genomic coordinates of mouse chromosome 12. The VH8‐8 gene with 500 bp of its upstream and downstream sequences (indicated by violet overlay) was inserted with the Floxin method at the deletion point of the Igh ∆890 allele to generate the Igh V8‐8 allele (Hill et al, 2020).
Comparison of the VDJ‐seq data obtained with pro‐B cells from the bone marrow of Igh +/+ (black) and Igh V8‐8/V8‐8 (violet) mice. The relative frequency of each VH gene is shown as a mean value with SEM and is based on five (Igh +/+) or three (Igh V8‐8/V8‐8) independent VDJ‐seq experiments. The different VH genes (horizontal axis) are aligned according to their position in the Igh locus (Dataset EV1). For further explanation, see legend of Fig 3D.
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) determined in pro‐B cells from Igh V8‐8/V8‐8 (violet), Igh +/+ (black), Igh ∆890/∆890 (white bar) and Igh P4/P4 (gray) mice. The average recombination frequency of the six distal VH genes was calculated as a mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1. Statistical data were analyzed by one‐way ANOVA (Tukey post hoc test); *P < 0.05, ***P < 0.001, ****P < 0.0001. Each dot corresponds to a mouse.
- A
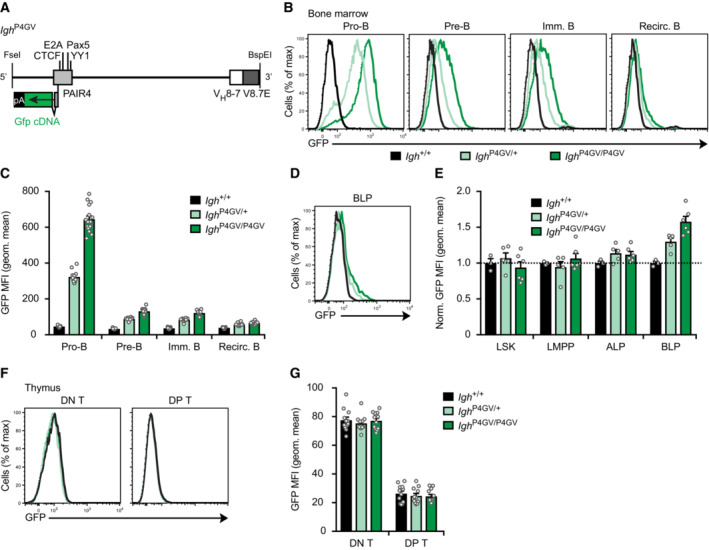
Schematic diagram of the Gfp‐linked PAIR4‐V8.7E module. The Gfp cDNA linked to 6 polyadenylation sites (pA) was inserted in exon 2 of PAIR4 prior to the insertion of the entire module at the deletion point into the Igh ∆890 allele by the Floxin method.
- B, C
Flow‐cytometric analysis (B) and geometric mean fluorescence intensity measurements (MFI, C) of GFP expression are shown for pro‐B, pre‐B, immature B, and recirculating B cells from the bone marrow of Igh P4GV/P4GV (dark green, n = 16), Igh P4GV/+ (light green, n = 12), and control Igh +/+ (black, n = 13) mice. Geometric mean fluorescence intensity measurements (C) are shown as mean values with SEM.
- D, E
Induction of GFP expression in a small subset of BLPs, as shown by flow‐cytometric analysis (D) and geometric mean fluorescence intensity measurements (E) of GFP expression in MPPs (LSK), LMPPs, ALPs, and BLPs from the bone marrow of the indicated mice. The dashed line indicates the absence of GFP expression in Igh +/+ cells, which were set to 1 for normalization of the MFI data. Geometric mean fluorescence intensity measurements (E) are shown as mean value with SEM based on 3 (Igh +/+), 5 (Igh P4GV/+) or 6 (Igh P4GV/P4GV) independent replicates.
- F, G
Absence of GFP expression in DN (CD4−CD8−) and DP (CD4+CD8+) T cells, as shown by flow‐cytometric analysis (F) and geometric mean fluorescence intensity measurements (G) of GFP expression in thymocytes from the indicated mice. Geometric mean fluorescence intensity measurements (G) are shown as a mean value with SEM based on 12 (Igh +/+), 11 (Igh P4GV/+), or 12 (Igh P4GV/P4GV) independent replicates.
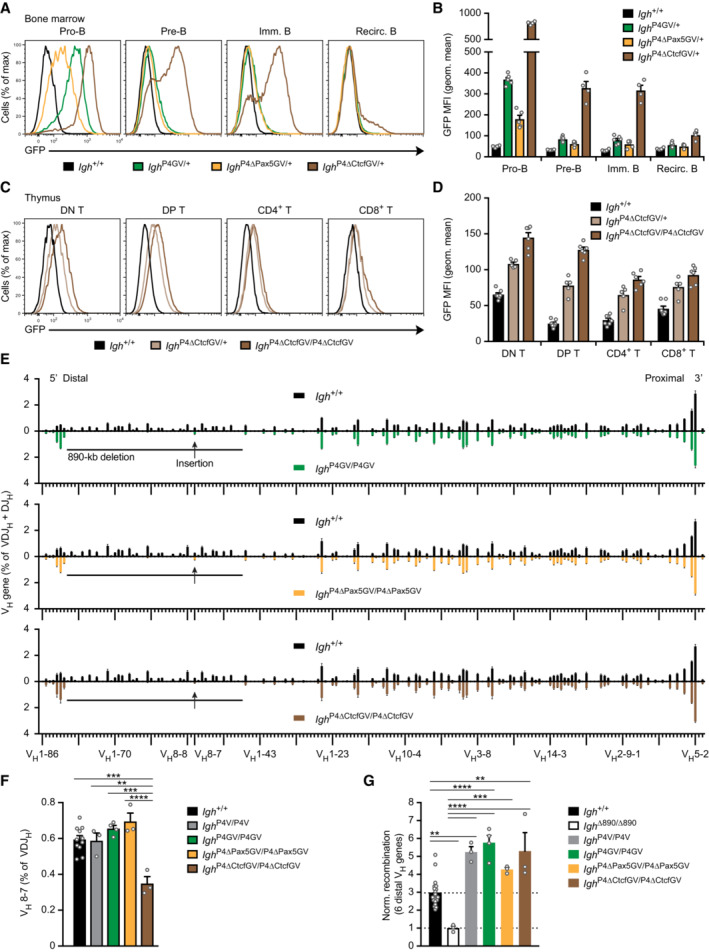
- A, B
Flow‐cytometric analysis (A) and geometric mean fluorescence intensity measurements (MFI, B) of GFP expression are shown for pro‐B, pre‐B, immature B, and recirculating B cells from the bone marrow of Igh P4GV/+ (green, n = 6), Igh P4∆Pax5GV/+ (yellow, n = 5), Igh P4∆CtcfGV/+ (brown, n = 4), and control Igh +/+ (black, n = 4) mice. Geometric mean fluorescence intensity measurements (B) are shown as a mean value with SEM.
- C, D
Activation of the PAIR4‐V8.7E module in thymocytes upon loss of CTCF binding at PAIR4, as shown by flow‐cytometric analysis (C) and geometric mean fluorescence intensity measurements (MFI, D) of GFP expression in DN, DP, CD4+, and CD8+ thymocytes of Igh P4∆CtcfGV/+ (light brown, n = 5) and Igh P4∆CtcfGV/P4∆CtcfGV (dark brown, n = 6) mice compared with control Igh +/+ (black, n = 7) mice. Geometric mean fluorescence intensity measurements (D) are shown as a mean value with SEM.
- E
Comparison of the VDJ‐seq data obtained with pro‐B cells of the Igh +/+ (black), Igh P4GV/P4GV (green), Igh P4∆Pax5GV/P4∆Pax5GV (yellow), and Igh P4∆CtcfGV/P4∆CtcfGV (brown) genotypes. The relative frequency of each VH gene is shown as mean value with SEM and is based on four (Igh +/+, Igh P4GV/P4GV) or three (Igh P4∆Pax5GV/P4∆Pax5GV, Igh P4∆CtcfGV/P4∆CtcfGV) independent VDJ‐seq experiments. The different VH genes (horizontal axis) are aligned according to their position in the Igh locus (Dataset EV1). For further explanation, see legend of Fig 3D.
- F
Recombination frequency of the inserted VH8‐7 gene, which was determined as percentage of all VDJH recombination events in pro‐B cells of the indicated genotypes and is shown as a mean value with SEM.
- G
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) determined in pro‐B cells of the indicated genotypes, based on the data shown in (E). The average recombination frequency of the six distal VH genes was calculated as mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1.
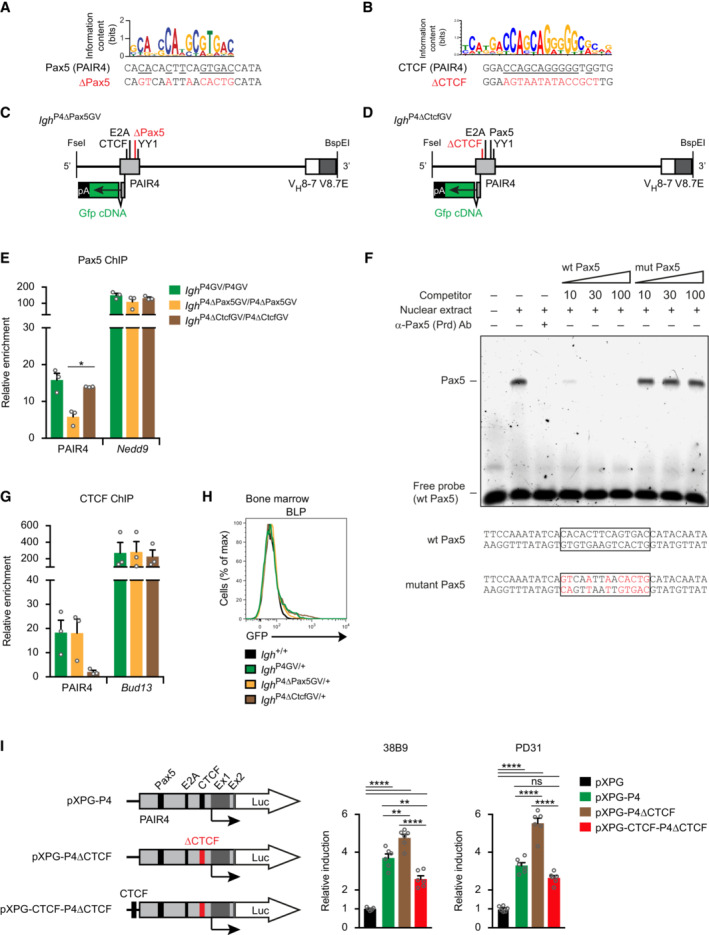
- A
Mutation of the Pax5‐binding sequence of PAIR4. The nucleotides of PAIR4, which match the Pax5 consensus recognition sequence (Kaiser et al, 2022), are underlined and were mutated to the nucleotides indicated in red (∆Pax5).
- B
Mutation of the CTCF‐binding sequence of PAIR4. The nucleotides of PAIR4, corresponding to the consensus CTCF‐binding motif (Hill et al, 2020), are underlined and were mutated to the nucleotides indicated in red (∆CTCF).
- C, D
Schematic diagram of the Gfp‐linked PAIR4‐V8.7E module with the Pax5‐ or CTCF‐binding site mutation, which was used for the generation of the Igh P4∆Pax5GV or Igh P4∆CtcfGV allele, respectively.
- E, G
Pax5 and CTCF binding to the wild‐type and mutant PAIR4 sequences. Short‐term cultured pro‐B cells from the bone marrow of Igh P4GV/P4GV (green), Igh P4∆Pax5GV/P4∆Pax5GV (yellow), and Igh P4∆CtcfGV/P4∆CtcfGV (brown) mice were used for ChIP analysis with an anti‐Pax5 paired domain antibody or an anti‐CTCF antibody. Input and precipitated DNA were quantified by qPCR analysis with primers amplifying the Pax5‐binding site present in PAIR4 or the Nedd9 gene (E), the CTCF‐binding site present in PAIR4 or the Bud13 gene (G) or by amplifying a gene‐poor region on chromosome 1 as a negative control (Dataset EV2). The amount of precipitated DNA was calculated as an average percentage of input with SEM and is shown relative to the value obtained for the negative control.
- F
No binding of Pax5 to the mutant Pax5 recognition sequence (∆Pax5) of PAIR4. A fluorescently‐labeled double‐stranded oligonucleotide containing the wild‐type (wt) Pax5 recognition sequence of PAIR4 (shown below) was used as a probe for electrophoretic mobility shift assay (EMSA) with a nuclear extract prepared from B cells of the human Ramos cell line (see Materials and Methods). The Pax5‐DNA complex (marked as Pax5 to the left) was not formed upon addition of an anti‐Pax5 antibody, directed against the DNA‐binding paired domain (Prd), or in the presence of a 10‐, 30‐, and 100‐fold molar excess of a nonlabeled competitor oligonucleotide containing the wild‐type Pax5‐binding site in contrast to the competitor oligonucleotide containing the mutant (mut) Pax5 recognition sequence (shown below).
- H
Absence of GFP expression in BLPs from the bone marrow of Igh P4GV/+ (green), Igh P4∆Pax5GV/+ (yellow), Igh P4∆CtcfGV/+ (brown), and control Igh +/+ (black) mice, as shown by flow‐cytometric analysis.
- I
Transient transfection reporter assay. The PAIR4‐luciferase reporter genes (schematically shown to the left) were transfected together with the control vector pRL‐CMV into cells of the pro‐B cell line 38B9 (Alt et al, 1984) or pre‐B cells line PD31 (Lewis et al, 1982). After 24 h, luciferase activities were measured, normalized and displayed relative to the activity of the parental vector pXPG, containing a promoter‐less firefly luciferase gene, which was set to 1. The relative luciferase (luc) activities of six independent transfection experiments are shown as mean values with SEM. Ex1 and Ex2 refer to exons 1 and 2 of the PAIR antisense transcript.
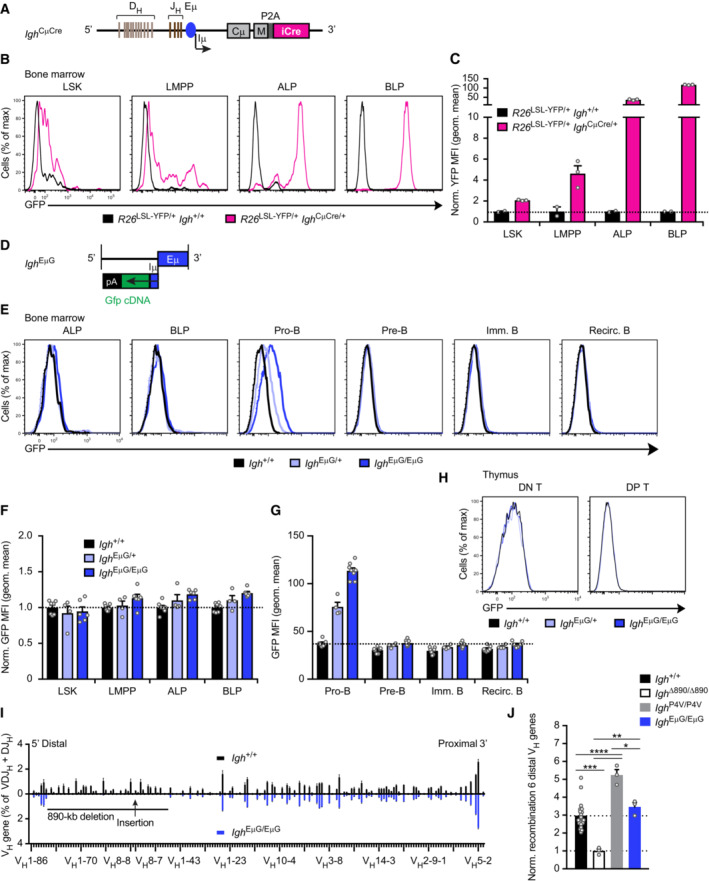
- A
Schematic diagram of the Igh CμCre allele, which contains the insertion of a Cre gene linked via a P2A sequence to the last codon of the second Cμ transmembrane exon (M2) in the Igh locus. The generation of the Igh CμCre allele is described in detail in Fig EV5A and Materials and Methods.
- B, C
Flow‐cytometric analysis (B) and geometric mean fluorescence intensity measurements (MFI, C) of YFP expression are shown for MPPs (LSK), LMPPs, ALPs, and BLPs from the bone marrow of Rosa26 LSL‐YPF/+ Igh CμCre/+ (violet) and control Rosa26 LSL‐YPF/+ Igh +/+ (black) mice. Geometric mean fluorescence intensity measurements (B) are shown as a mean value with SEM and have been normalized by setting the mean value obtained with Igh +/+ cells to 1.
- D
Schematic diagram of the Gfp‐linked Iμ‐Eμ sequences used to generate the Igh EμG allele. The Gfp cDNA together with six polyadenylation sites (pA) was linked to the Iμ exon downstream of the Eμ enhancer prior to insertion of these sequence at the deletion point into the Igh ∆890 allele by the Floxin method.
- E–G
Flow‐cytometric analysis (E) and geometric mean fluorescence intensity measurements (MFI, F, and G) of GFP expression are shown for MPPs (LSK), LMPPs, ALPs, BLPs, pro‐B, pre‐B, immature B, and recirculating B cells from the bone marrow of Igh EμG/+ (light blue), Igh EμG/EμG (dark blue), and control Igh +/+ (black) mice. Geometric mean fluorescence intensity measurements (B) are shown as mean a value with SEM. For lymphoid progenitors, MFI values have been normalized by setting the mean value obtained with Igh +/+ cells to 1. The flow‐cytometric definition of the lymphoid progenitors, B and T cell types is described in Materials and Methods.
- H
Absence of GFP expression in DN and DP T cells from the thymus of Igh EμG/+ (light blue) and Igh EμG/EμG (dark blue) mice compared with control Igh +/+ (black) mice.
- I
Comparison of the VDJ‐seq data obtained with pro‐B cells of the Igh +/+ (black) and Igh EμG/EμG (dark blue) genotypes. The relative frequency of each VH gene is shown as mean value with SEM and is based on six (Igh +/+) or three (Igh EμG/EμG) independent VDJ‐seq experiments. For further description, see legend of Fig 3D.
- J
Normalized recombination frequency of the first six distal VH genes (VH1‐85 to VH1‐80) determined in pro‐B cells of the indicated genotypes, based on the data shown in (I). The average recombination frequency of the six distal VH genes was calculated as a mean value with SEM, and the value obtained with Igh ∆890/∆890 pro‐B cells was set to 1.
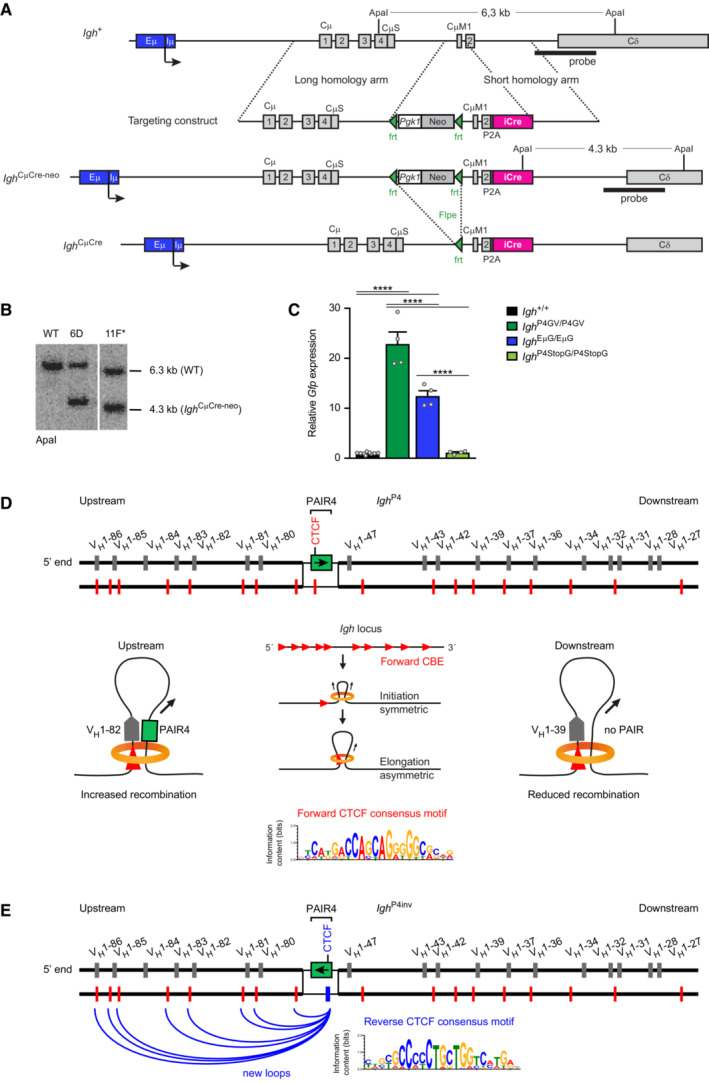
Generation of the Igh CμCre allele by ES cell targeting. The Igh CμCre‐neo allele was generated by homologous recombination in the ES cell line A9 by using the following targeting vector. The targeting vector consisted of a 4.1‐kb long 5′ homology region (containing Cμ1 to CμS), a frt‐flanked 2.1‐kb DNA fragment containing the mouse phosphoglycerate kinase (Pgk1) promoter linked to the neomycin (Neo) resistance gene and a SV40 polyadenylation signal, a 1.6‐kb DNA fragment containing the CμM1/2 sequence linked in frame via the P2A peptide to an iCre gene followed by a 1.8‐kb long 3′ homology region. The frt sites are indicated by green arrowheads. The ApaI fragments of the Igh + and Igh CμCre‐neo alleles, which were used for allele identification by Southern blot analysis with the indicated probe, are shown together with their length (in kilobases, kb). The Igh CμCre allele was generated by deletion of the frt‐flanked Pgk1‐Neo expression cassette, which was used for selection of the targeted ES cell clones, in Igh CμCre‐neo mice expressing the Flpe transgene.
Southern blot analysis of wild‐type (WT) ES cells and correctly targeted ES cell clones by hybridization of ApaI‐digested genomic DNA with the probe indicated in (A). The ES cell clone 11F was injected into blastocysts to generate the Igh CμCre‐neo/+ mouse strain.
RT‐qPCR analysis of Gfp mRNA expression in sorted pro‐B cells from the bone marrow of the indicated mouse strains. The transcripts of the Gfp gene were normalized against the control Tbp mRNA, and the mean value obtained with Igh +/+ pro‐B cells was set to 1. Gfp mRNA expression is shown as a mean value with SEM based on nine (Igh +/+) or four (Igh P4GV/P4GV, Igh EμG/EμG, Igh P4StopG/P4StopG) independent samples. Statistical data were analyzed by one‐way ANOVA (Tukey post hoc test); ****P < 0.0001.
Role of loop extrusion in mediating increased recombination of VH genes located upstream of the insertion of the PAIR4‐V8.7E module (Figs 3G and EV3D). The loop extrusion process has been shown to generate a largely contiguous interaction zone where all the different sequences within the VH gene region appear to interact with each other (Hill et al, 2023). The generation of this interaction zone requires that all CTCF‐binding sites are present in forward orientation in the VH gene cluster (Hill et al, 2020). Loop extrusion likely initiates at random positions in the VH gene cluster and initially proceeds in a symmetrical manner until the cohesin ring interacts with a CTCF protein bound to the next upstream forward‐oriented CTCF‐binding site, which leads to stabilized binding of cohesin at this site (Li et al, 2020). Thereafter, asymmetrical loop extrusion reels the DNA of the downstream Igh regions into the loop, until it is halted by a CTCF protein bound to a reverse‐oriented CTCF‐binding site at the IGCR1 or 3′CBE elements (Hill et al, 2023). This loop extrusion mechanism predicts that only the VH1 genes located upstream of the PAIR4 insertion can efficiently interact through loop extrusion with the inserted PAIR4 element and thus undergo increased VH1 gene recombination. In contrast, the downstream VH1 genes are unable to interact through this loop extrusion mechanism with the inserted PAIR4 element, which results in reduced VH1 gene recombination.
Explanation for the reduced recombination efficiency of upstream VH1 genes in Igh P4inv/P4inv pro‐B cells compared with Igh P4/P4 pro‐B cells (Fig 3G and F). The inversion of PAIR4 also inverts the orientation of the CTCF‐binding site of PAIR4 in Igh P4inv/P4inv pro‐B cells relative to Igh P4/P4 pro‐B cells. The reverse‐oriented CTCF‐binding site of PAIR4 is now in convergent orientation relative to the forward CTCF‐binding sites in the upstream region and can thus form new stabilized loops (Rao et al, 2014) that interfere with prolonged loop extrusion beyond the inserted PAIR4 element and can therefore prevent interactions of the upstream VH genes with the RAG+ recombination center in the Igh 3′ region, as previously shown for the inversion of the 890‐kb VH gene region (Hill et al, 2020). The forward and reverse CTCF consensus motifs (D, E) are shown.
Similar articles
-
Contribution of the IGCR1 regulatory element and the 3'Igh CTCF-binding elements to regulation of Igh V(D)J recombination.Proc Natl Acad Sci U S A. 2023 Jun 27;120(26):e2306564120. doi: 10.1073/pnas.2306564120. Epub 2023 Jun 20. Proc Natl Acad Sci U S A. 2023. PMID: 37339228 Free PMC article.
-
Epigenetic modifications of the VH region after DJH recombination in Pro-B cells.Immunology. 2017 Oct;152(2):218-231. doi: 10.1111/imm.12758. Epub 2017 Jun 23. Immunology. 2017. PMID: 28502113 Free PMC article.
-
Pax5 induces V-to-DJ rearrangements and locus contraction of the immunoglobulin heavy-chain gene.Genes Dev. 2004 Feb 15;18(4):411-22. doi: 10.1101/gad.291504. Genes Dev. 2004. PMID: 15004008 Free PMC article.
-
Spatial Regulation of V-(D)J Recombination at Antigen Receptor Loci.Adv Immunol. 2015;128:93-121. doi: 10.1016/bs.ai.2015.07.006. Epub 2015 Aug 13. Adv Immunol. 2015. PMID: 26477366 Review.
-
Epigenetic Regulation of Individual Modules of the immunoglobulin heavy chain locus 3' Regulatory Region.Front Immunol. 2014 Apr 21;5:163. doi: 10.3389/fimmu.2014.00163. eCollection 2014. Front Immunol. 2014. PMID: 24795714 Free PMC article. Review.
Cited by
-
Dynamic O-GlcNAcylation governs long-range chromatin interactions in V(D)J recombination during early B-cell development.Cell Mol Immunol. 2025 Jan;22(1):68-82. doi: 10.1038/s41423-024-01236-9. Epub 2024 Dec 3. Cell Mol Immunol. 2025. PMID: 39627609
References
-
- Adams B, Dörfler P, Aguzzi A, Kozmik Z, Urbánek P, Maurer‐Fogy I, Busslinger M (1992) Pax‐5 encodes the transcription factor BSAP and is expressed in B lymphocytes, the developing CNS, and adult testis. Genes Dev 6: 1589–1607 - PubMed
-
- Anastassiadis K, Fu J, Patsch C, Hu S, Weidlich S, Duerschke K, Buchholz F, Edenhofer F, Stewart AF (2009) Dre recombinase, like Cre, is a highly efficient site‐specific recombinase in E. coli, mammalian cells and mice. Dis Model Mech 2: 508–515 - PubMed
-
- Baniahmad A, Steiner C, Kohne AC, Renkawitz R (1990) Modular structure of a chicken lysozyme silencer: involvement of an unusual thyroid hormone receptor binding site. Cell 61: 505–514 - PubMed
Publication types
MeSH terms
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
