

Molecular and cellular evidence for the impact of a hypertrophic cardiomyopathy-associated RAF1 variant on the structure and function of contractile machinery in bioartificial cardiac tissues
- PMID: 37344639
- PMCID: PMC10284840
- DOI: 10.1038/s42003-023-05013-8
Molecular and cellular evidence for the impact of a hypertrophic cardiomyopathy-associated RAF1 variant on the structure and function of contractile machinery in bioartificial cardiac tissues
Abstract
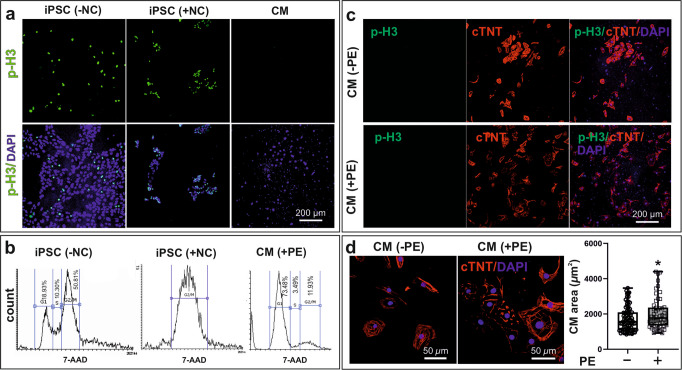
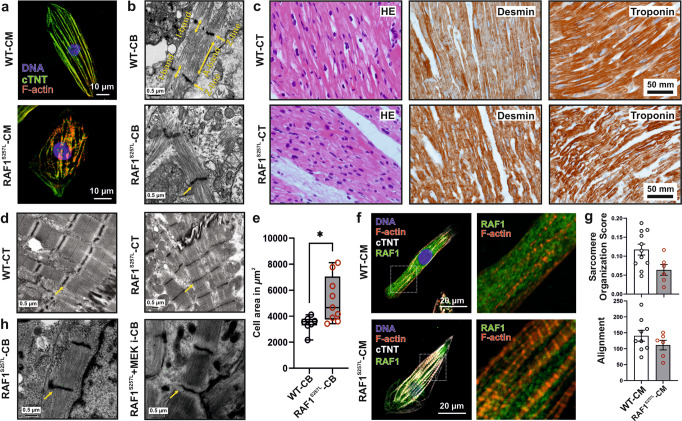
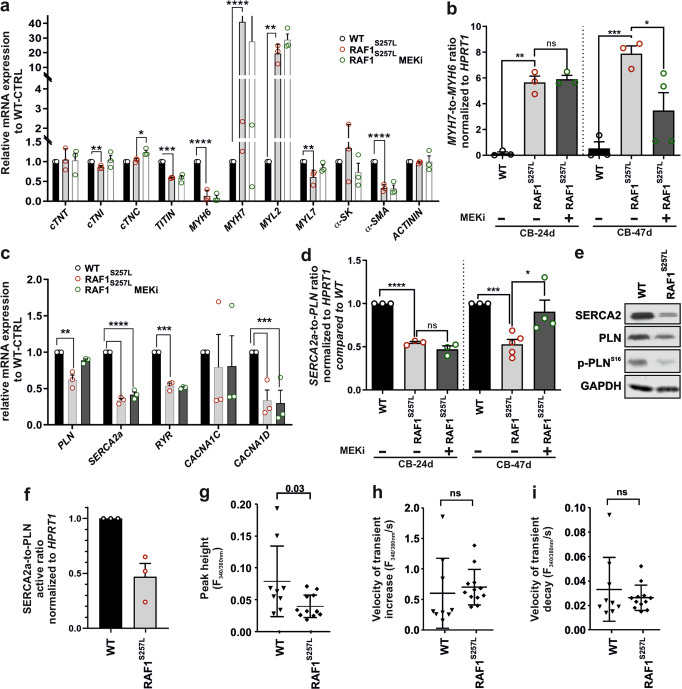
Noonan syndrome (NS), the most common among RASopathies, is caused by germline variants in genes encoding components of the RAS-MAPK pathway. Distinct variants, including the recurrent Ser257Leu substitution in RAF1, are associated with severe hypertrophic cardiomyopathy (HCM). Here, we investigated the elusive mechanistic link between NS-associated RAF1S257L and HCM using three-dimensional cardiac bodies and bioartificial cardiac tissues generated from patient-derived induced pluripotent stem cells (iPSCs) harboring the pathogenic RAF1 c.770 C > T missense change. We characterize the molecular, structural, and functional consequences of aberrant RAF1-associated signaling on the cardiac models. Ultrastructural assessment of the sarcomere revealed a shortening of the I-bands along the Z disc area in both iPSC-derived RAF1S257L cardiomyocytes and myocardial tissue biopsies. The aforementioned changes correlated with the isoform shift of titin from a longer (N2BA) to a shorter isoform (N2B) that also affected the active force generation and contractile tensions. The genotype-phenotype correlation was confirmed using cardiomyocyte progeny of an isogenic gene-corrected RAF1S257L-iPSC line and was mainly reversed by MEK inhibition. Collectively, our findings uncovered a direct link between a RASopathy gene variant and the abnormal sarcomere structure resulting in a cardiac dysfunction that remarkably recapitulates the human disease.
© 2023. The Author(s).
Conflict of interest statement
B.D.G.’s institution receives royalties from genetic testing for RAF1 for Noonan syndrome from Correlegan, LabCorp, GeneDx, and Prevention Genetics. B.D.G. is a consultant for Day One Biopharmaceuticals and was recently a consultant for BioMarin. He received a sponsored research award from Onconova. M.Z. is a member of a scientific advisory board for Day One Biopharmaceuticals.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
