

Wide-field fluorescence lifetime imaging of neuron spiking and subthreshold activity in vivo
- PMID: 37347862
- PMCID: PMC10361454
- DOI: 10.1126/science.adf9725
Wide-field fluorescence lifetime imaging of neuron spiking and subthreshold activity in vivo
Abstract
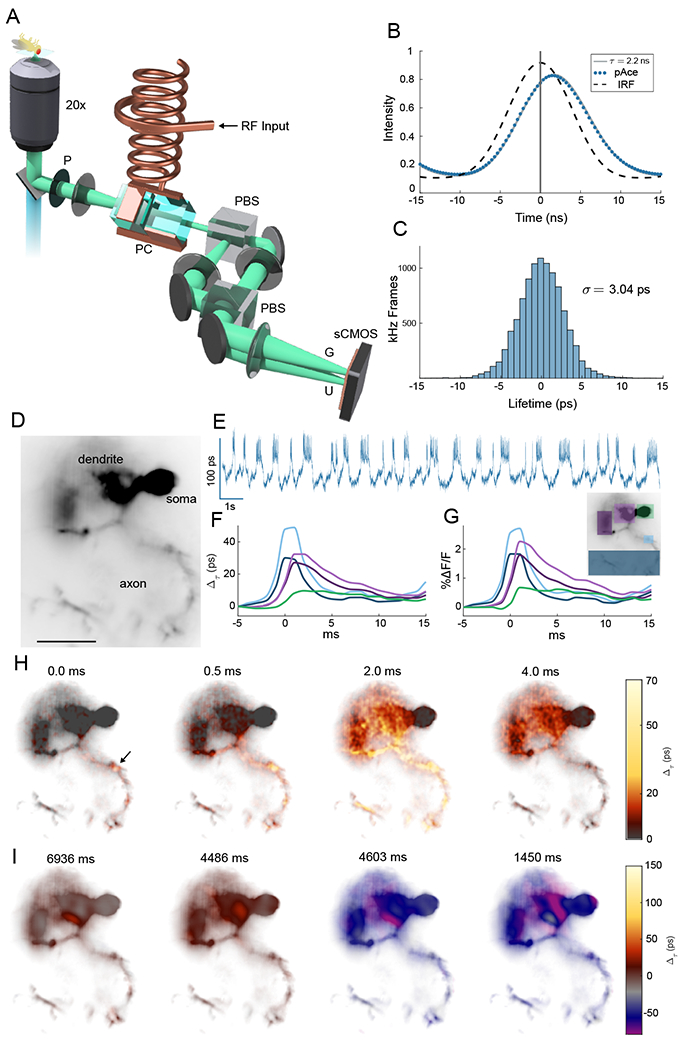
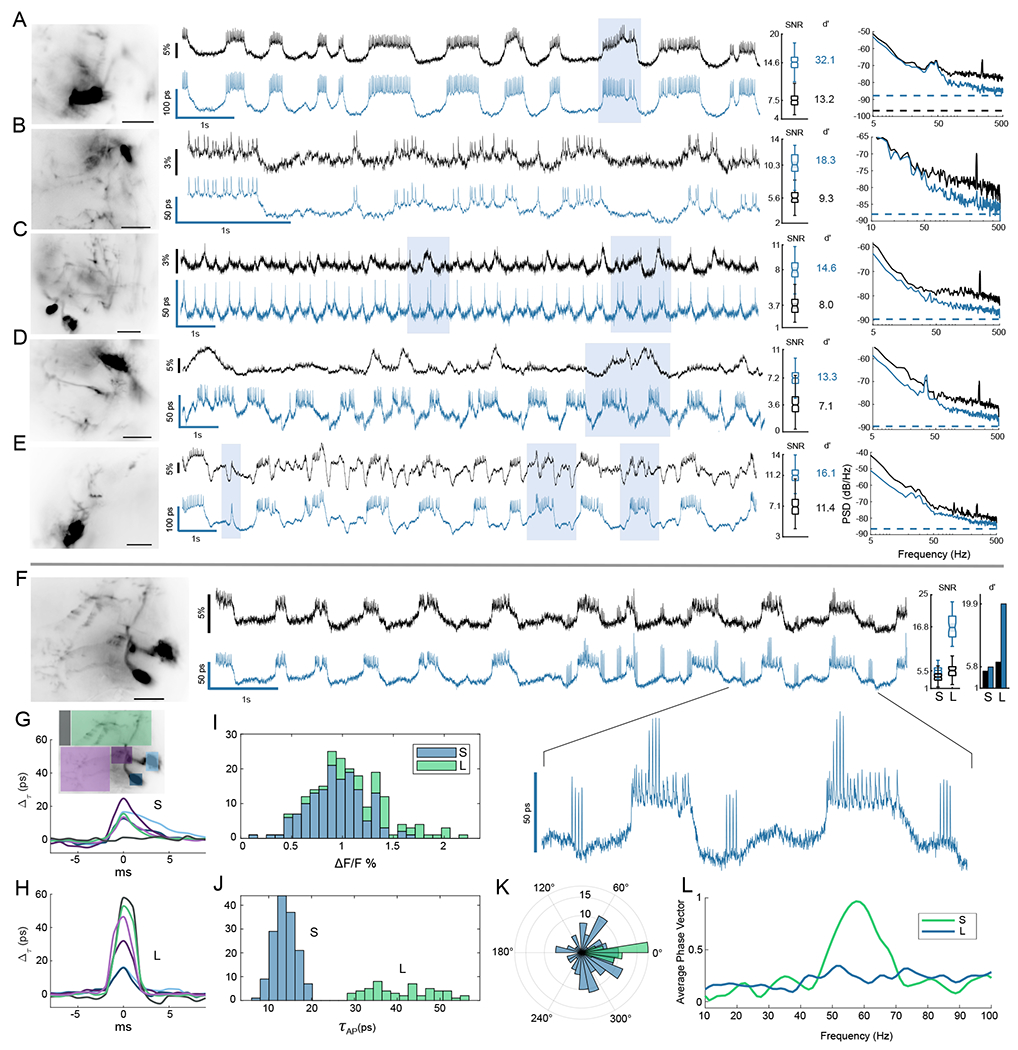
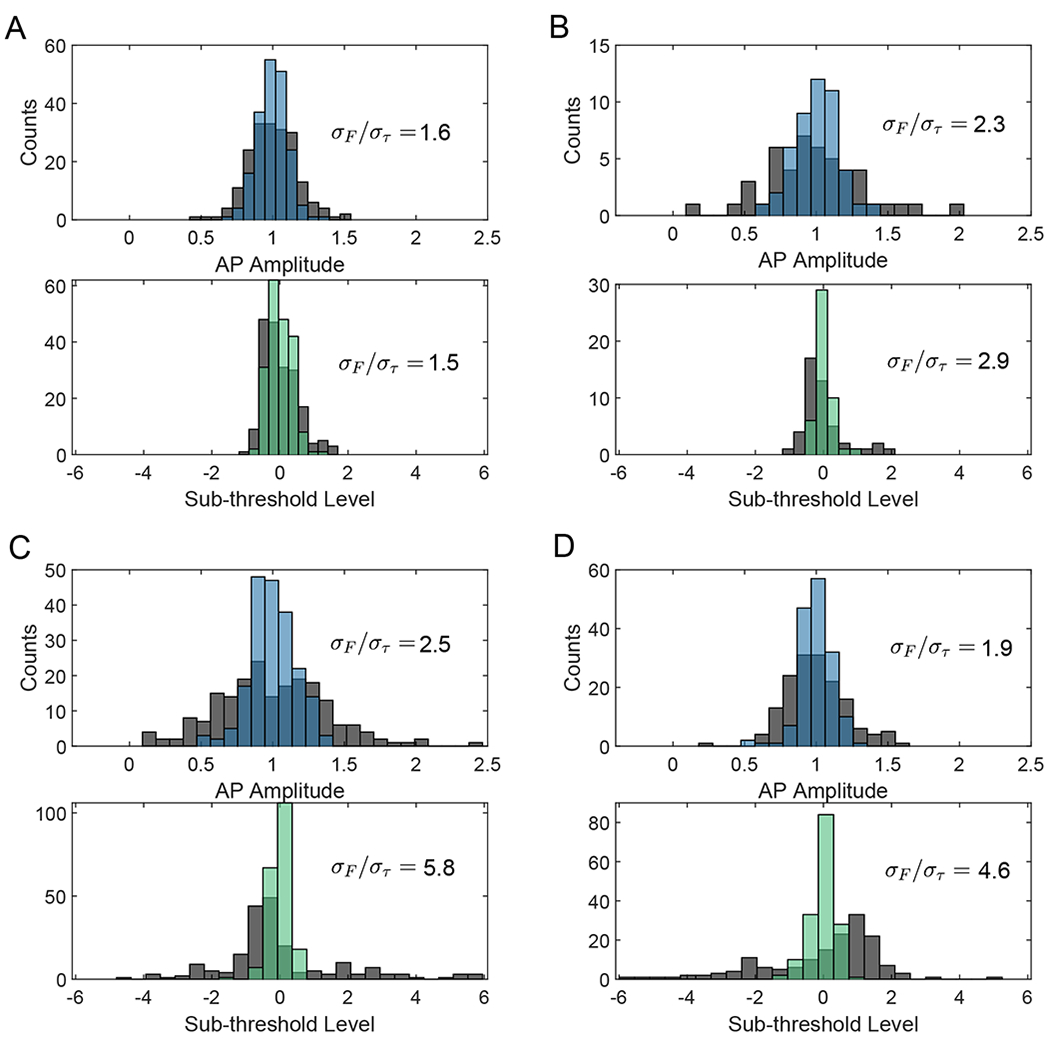
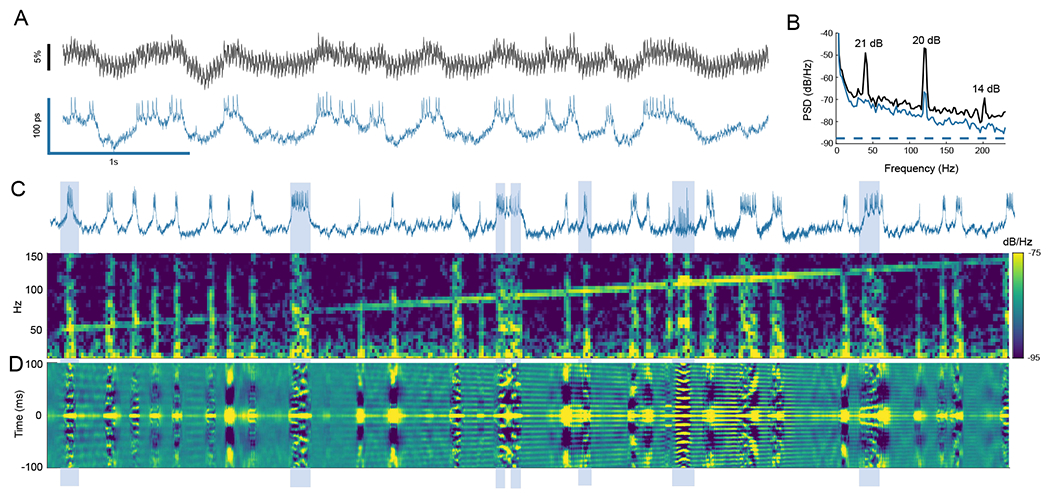
The development of voltage-sensitive fluorescent probes suggests fluorescence lifetime as a promising readout for electrical activity in biological systems. Existing approaches fail to achieve the speed and sensitivity required for voltage imaging in neuroscience applications. We demonstrated that wide-field electro-optic fluorescence lifetime imaging microscopy (EO-FLIM) allows lifetime imaging at kilohertz frame-acquisition rates, spatially resolving action potential propagation and subthreshold neural activity in live adult Drosophila. Lifetime resolutions of <5 picoseconds at 1 kilohertz were achieved for single-cell voltage recordings. Lifetime readout is limited by photon shot noise, and the method provides strong rejection of motion artifacts and technical noise sources. Recordings revealed local transmembrane depolarizations, two types of spikes with distinct fluorescence lifetimes, and phase locking of spikes to an external mechanical stimulus.
Conflict of interest statement
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
