

Host ranges of Sf-rhabdoviruses harbored by lepidopteran insects and insect cell lines
- PMID: 37348145
- PMCID: PMC10528406
- DOI: 10.1016/j.virol.2023.05.007
Host ranges of Sf-rhabdoviruses harbored by lepidopteran insects and insect cell lines
Abstract
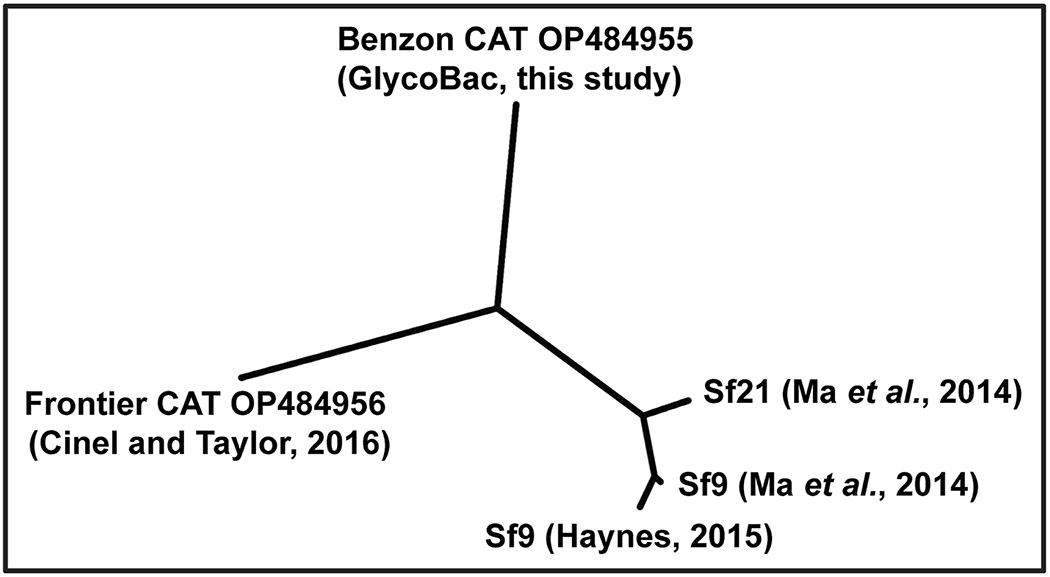
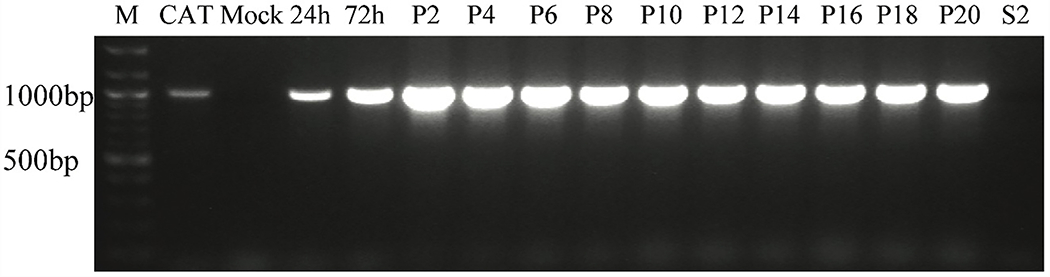
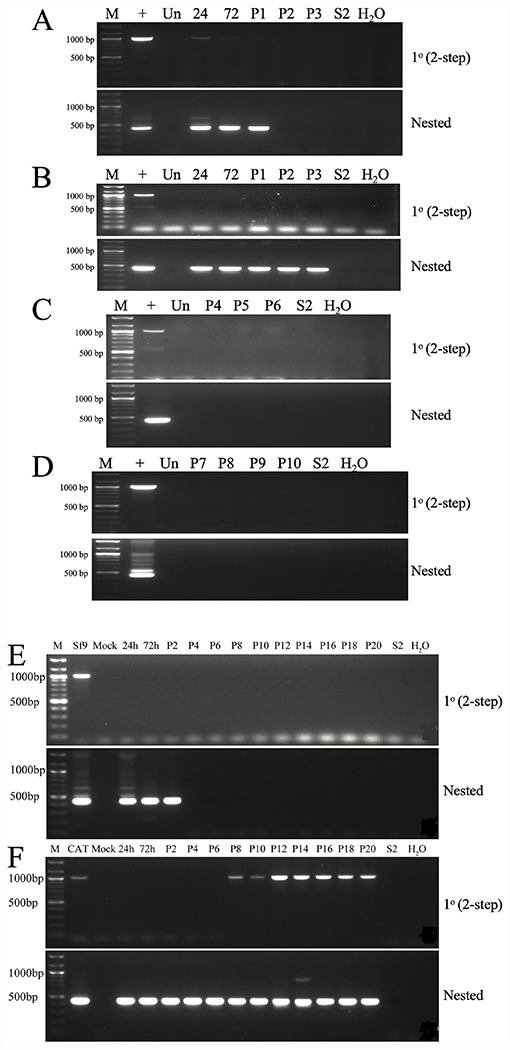
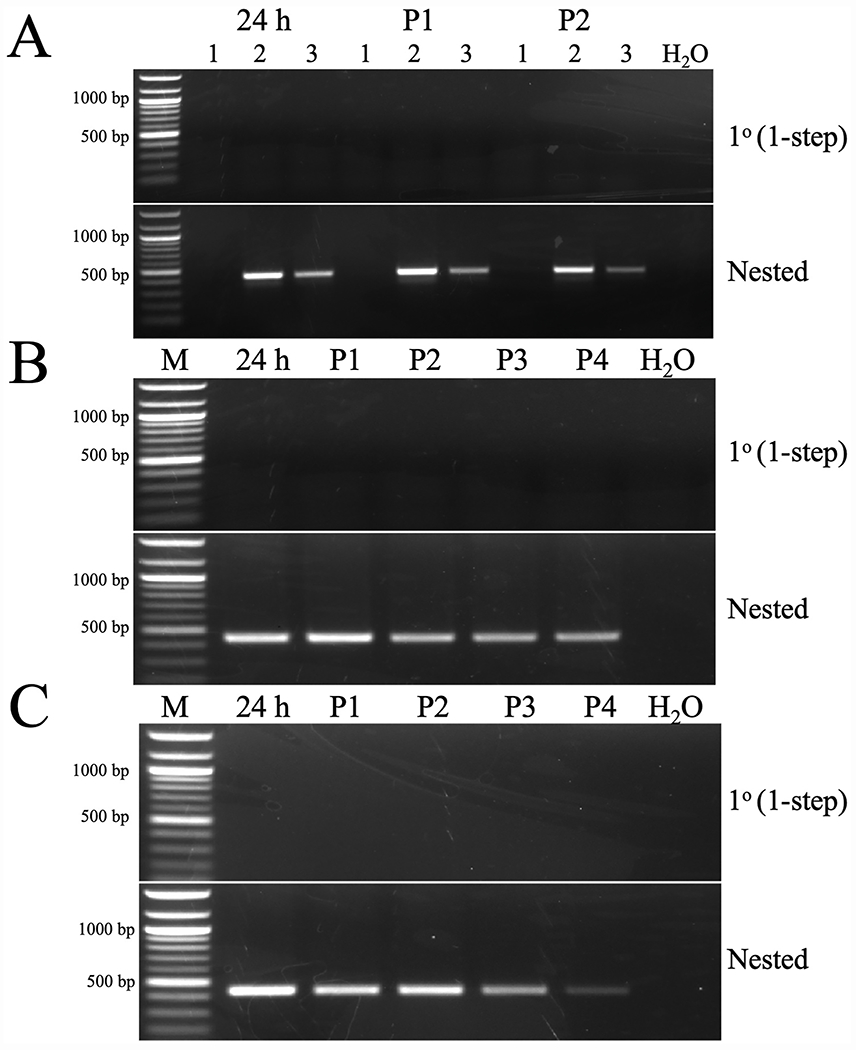
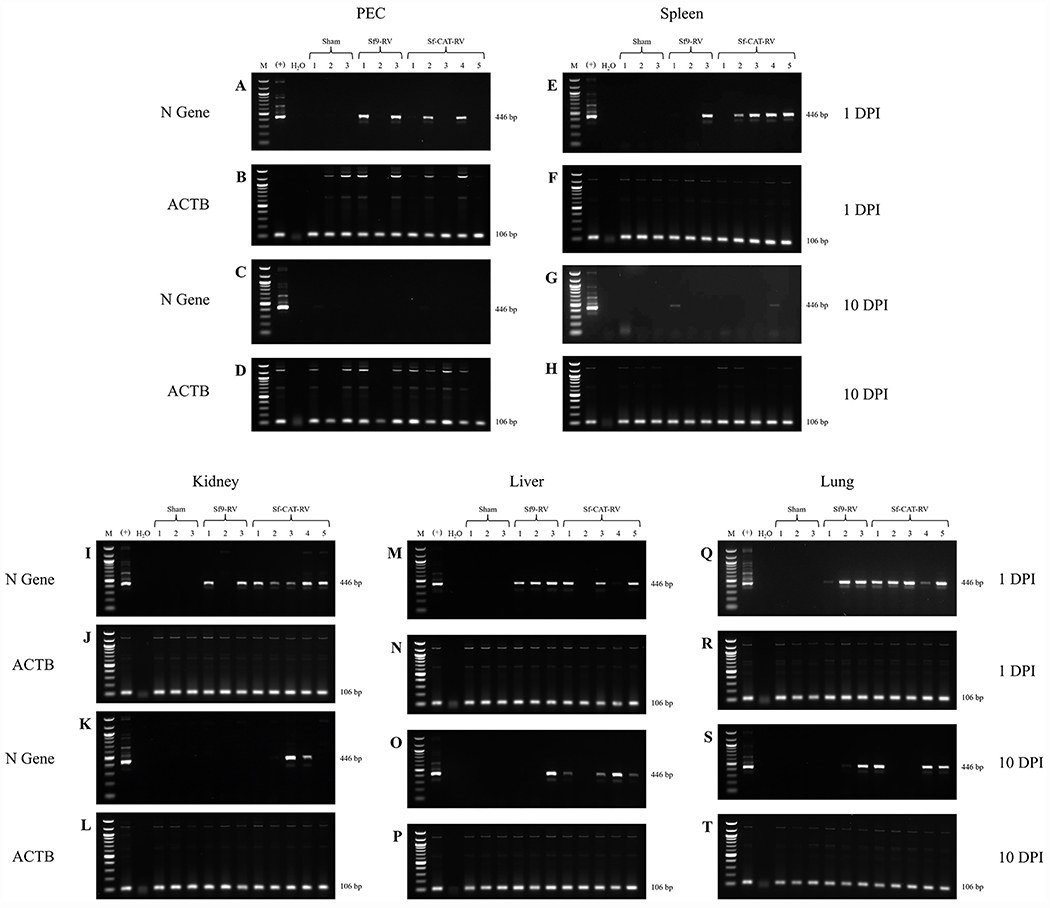
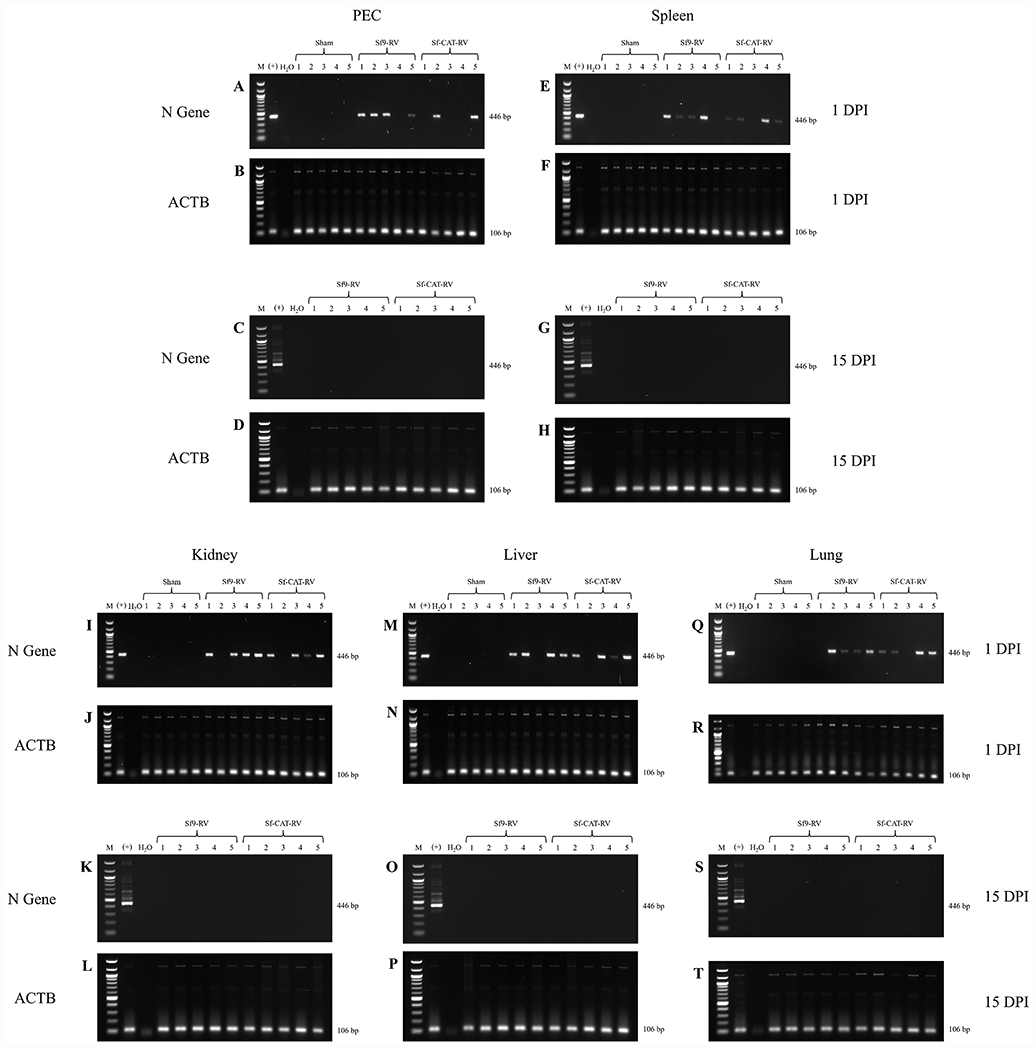
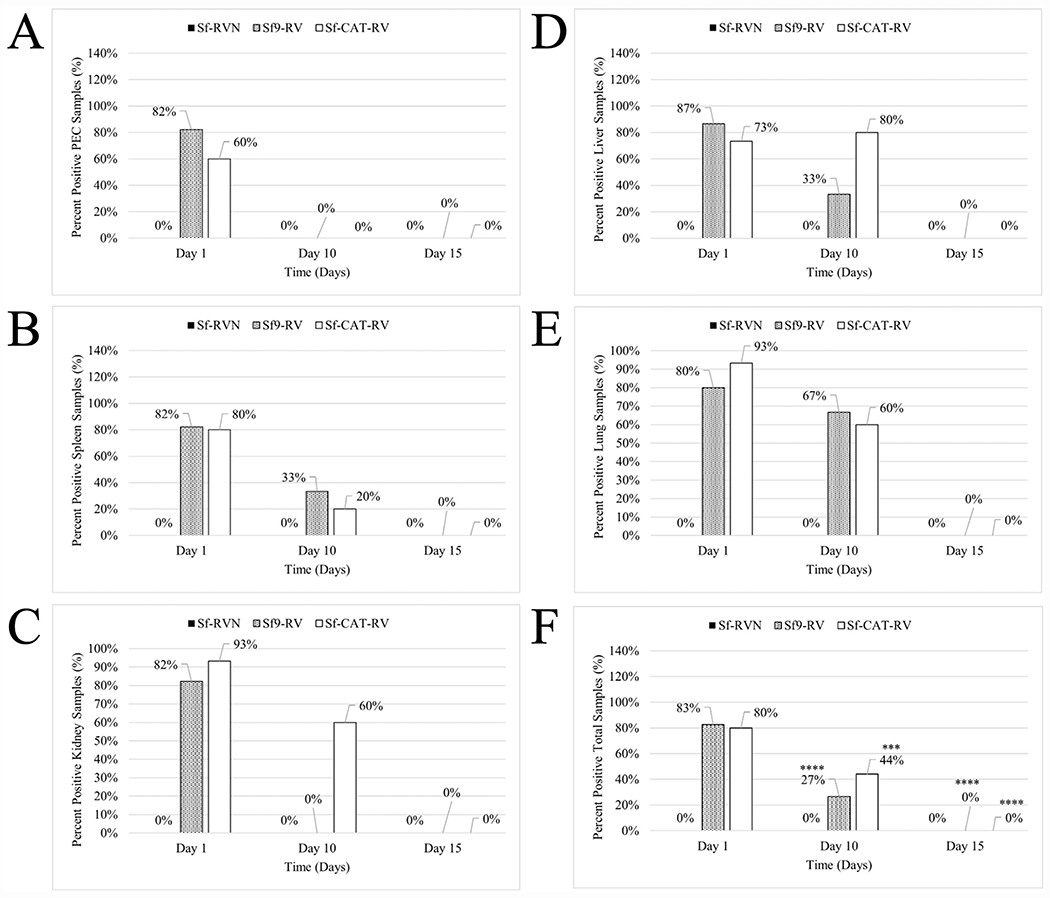
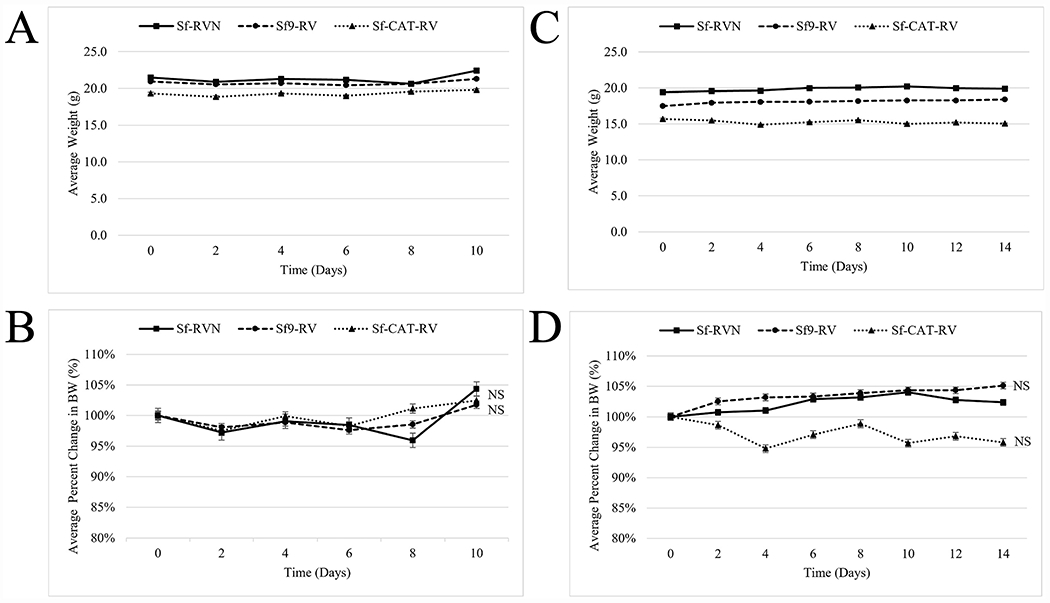
Cell lines derived from Spodoptera frugiperda (Sf), which are the most widely used hosts in the baculovirus-insect cell system, are contaminated with Sf-rhabdoviruses (Sf-RVs). In this study, we identified a closely related virus (Sf-CAT-RV) in the caterpillar species used to isolate the original Sf cell line. We then evaluated the Sf-RV and Sf-CAT-RV host ranges, found Sf-CAT-RV could infect Vero cells, and obtained results suggesting both variants can infect mouse ear fibroblasts. In addition, we found both variants could establish pantropic infections in severely immunocompromised (RAG2/IL2RG-/-) mice. However, both variants were cleared by two weeks post-inoculation and neither produced any symptoms or obvious adverse outcomes in these hosts. We conclude the caterpillars used to isolate Sf21 cells were the most likely source of the Sf-RV contaminant, Sf-RVs and their Sf-CAT-RV progenitor have broader host ranges than expected from previous work, but neither variant poses a serious threat to human health.
Keywords: Adventitious viral contamination; Baculovirus-insect cell expression system; Biosafety; Insect cell lines; Sf-rhabdovirus.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Host Range and Population Survey of Spodoptera frugiperda Rhabdovirus.J Virol. 2019 Mar 5;93(6):e02028-18. doi: 10.1128/JVI.02028-18. Print 2019 Mar 15. J Virol. 2019. PMID: 30626676 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
Comparison of Two Modern Survival Prediction Tools, SORG-MLA and METSSS, in Patients With Symptomatic Long-bone Metastases Who Underwent Local Treatment With Surgery Followed by Radiotherapy and With Radiotherapy Alone.Clin Orthop Relat Res. 2024 Dec 1;482(12):2193-2208. doi: 10.1097/CORR.0000000000003185. Epub 2024 Jul 23. Clin Orthop Relat Res. 2024. PMID: 39051924
-
Topical antiseptics for chronic suppurative otitis media.Cochrane Database Syst Rev. 2025 Jun 9;6(6):CD013055. doi: 10.1002/14651858.CD013055.pub3. Cochrane Database Syst Rev. 2025. PMID: 40484403
-
Inhaled steroids and risk of pneumonia for chronic obstructive pulmonary disease.Cochrane Database Syst Rev. 2014 Mar 10;2014(3):CD010115. doi: 10.1002/14651858.CD010115.pub2. Cochrane Database Syst Rev. 2014. PMID: 24615270 Free PMC article.
Cited by
-
A novel rhabdovirus persistently infects lepidopteran cell lines.Virus Res. 2025 Apr;354:199548. doi: 10.1016/j.virusres.2025.199548. Epub 2025 Feb 21. Virus Res. 2025. PMID: 39988207 Free PMC article.
References
-
- Food and Drug Administration, 2010. Guidance for Industry - Characterization and Qualification of Cell Substrates and Other Biological Materials Used in the Production of Viral Vaccines for Infectious Disease Indications, in: Research, C.f.B.E.a (Ed.). Office of Communication, Outreach and Development, Rockville, MD.
-
- Garcia S, DiSanto J, Stockinger B, 1999. Following the development of a CD4 T cell response in vivo: from activation to memory formation. Immunity 11, 163–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
