

GARP on hepatic stellate cells is essential for the development of liver fibrosis
- PMID: 37348791
- PMCID: PMC10592496
- DOI: 10.1016/j.jhep.2023.05.043
GARP on hepatic stellate cells is essential for the development of liver fibrosis
Abstract
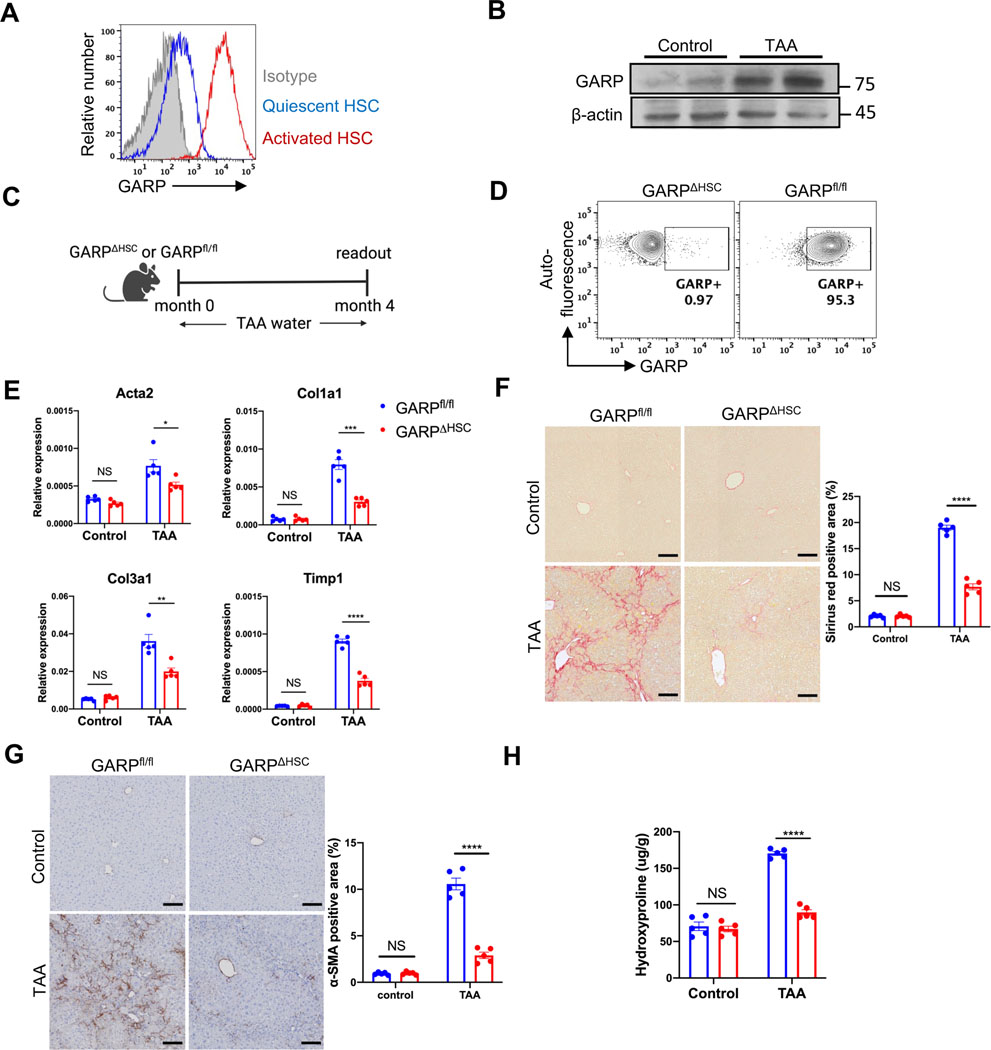
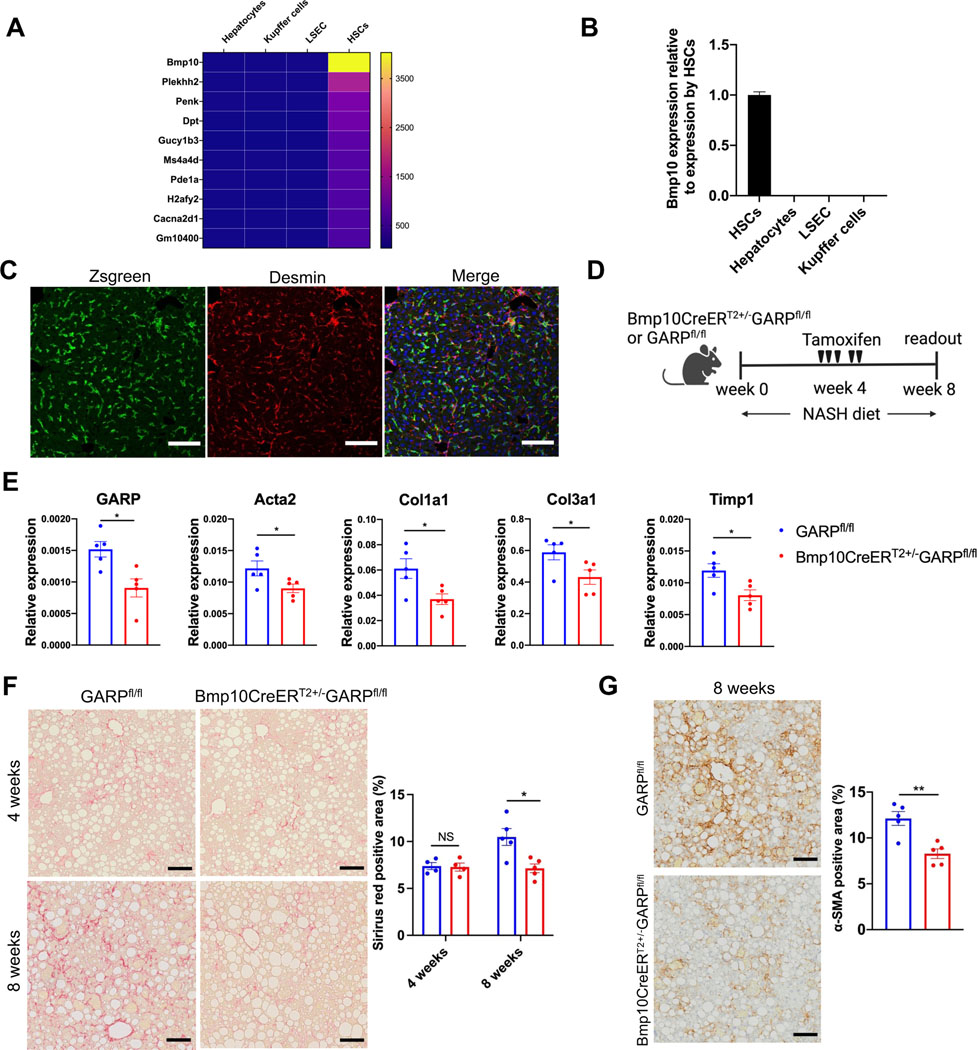
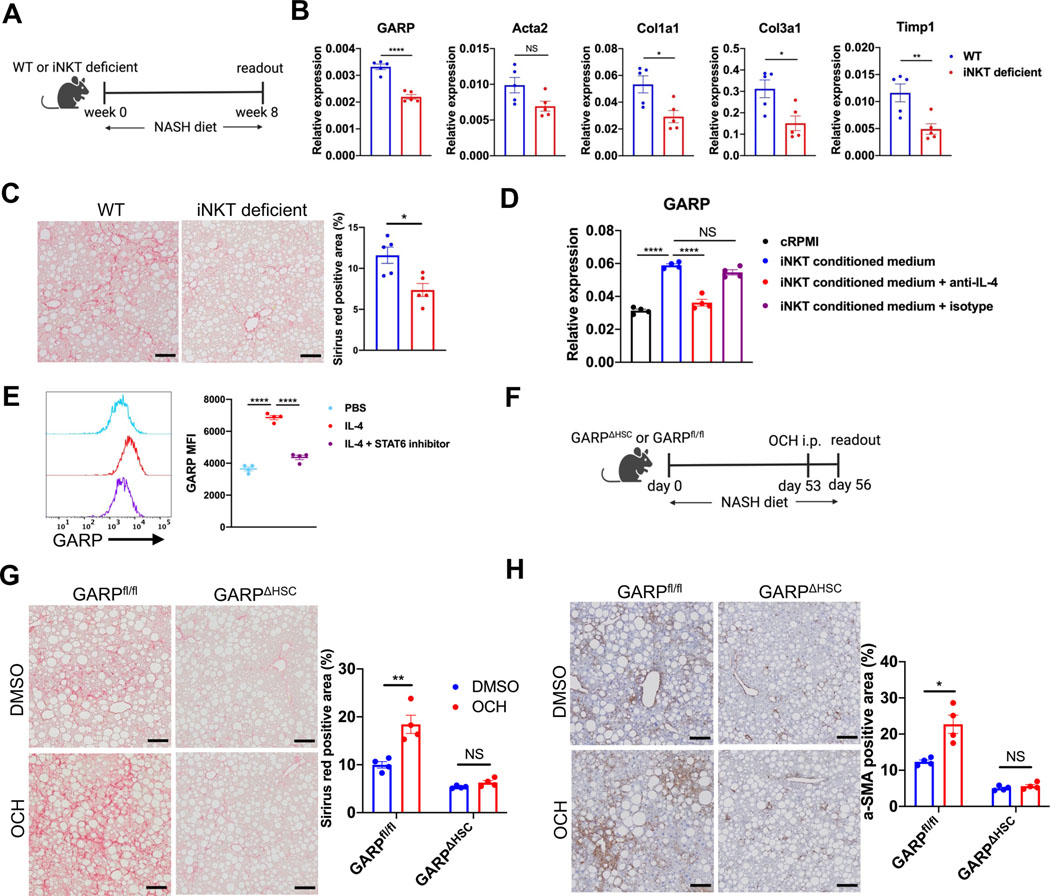
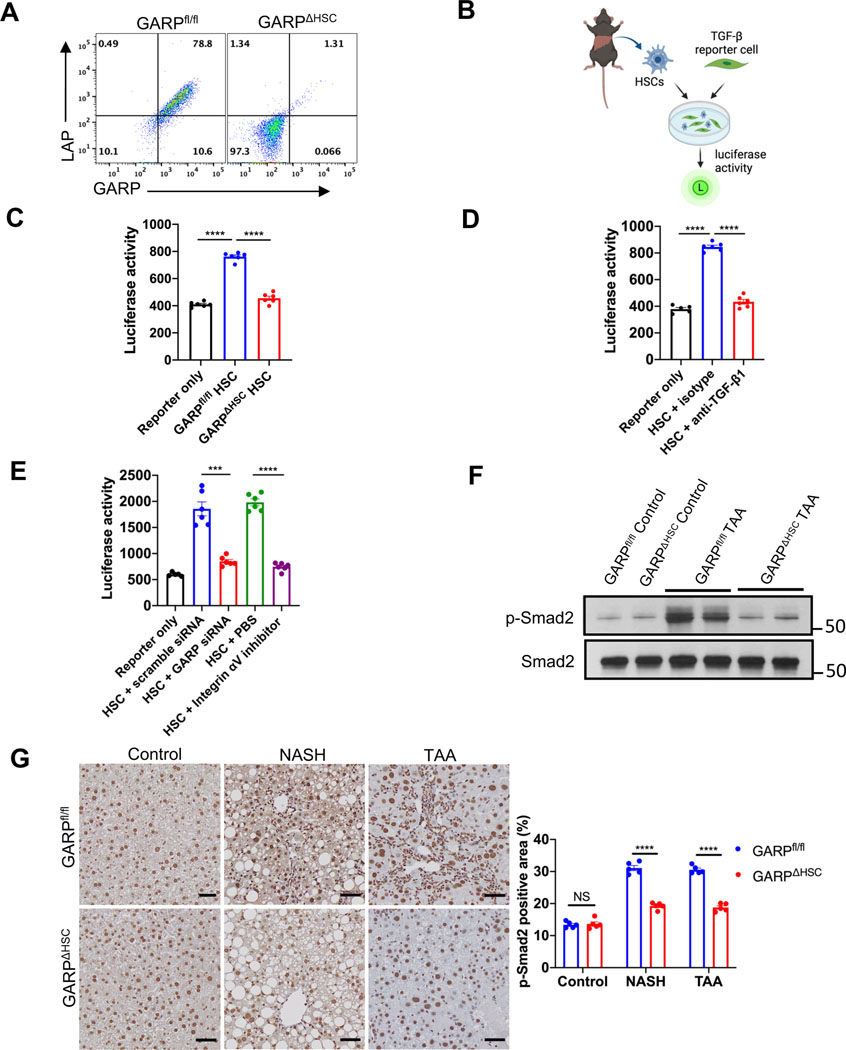
Background & aims: Glycoprotein A repetitions predominant (GARP) is a membrane protein that functions as a latent TGF-β docking molecule. While the immune regulatory properties of GARP on blood cells have been studied, the function of GARP on tissue stromal cells remains unclear. Here, we investigate the role of GARP expressed on hepatic stellate cells (HSCs) in the development of liver fibrosis.
Methods: The function of GARP on HSCs was explored in toxin-induced and metabolic liver fibrosis models, using conditional GARP-deficient mice or a newly generated inducible system for HSC-specific gene ablation. Primary mouse and human HSCs were isolated to evaluate the contribution of GARP to the activation of latent TGF-β. Moreover, cell contraction of HSCs in the context of TGF-β activation was tested in a GARP-dependent fashion.
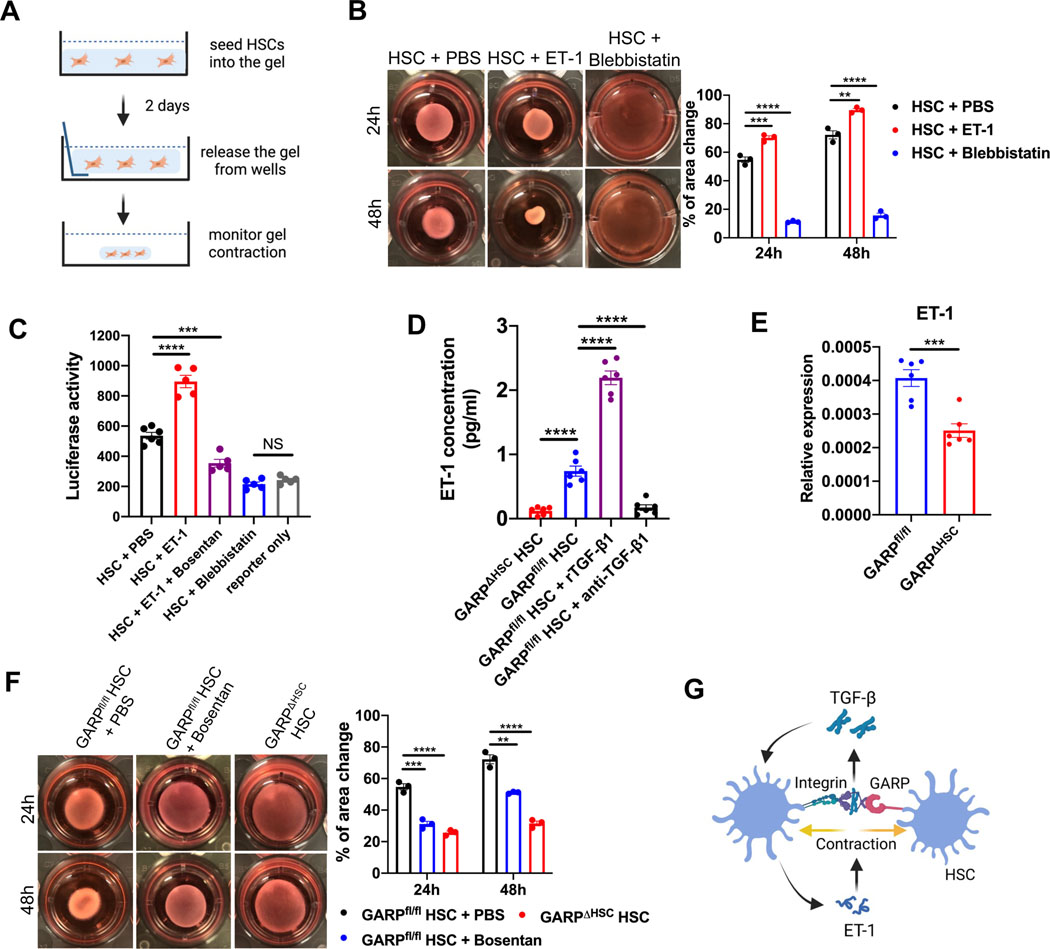
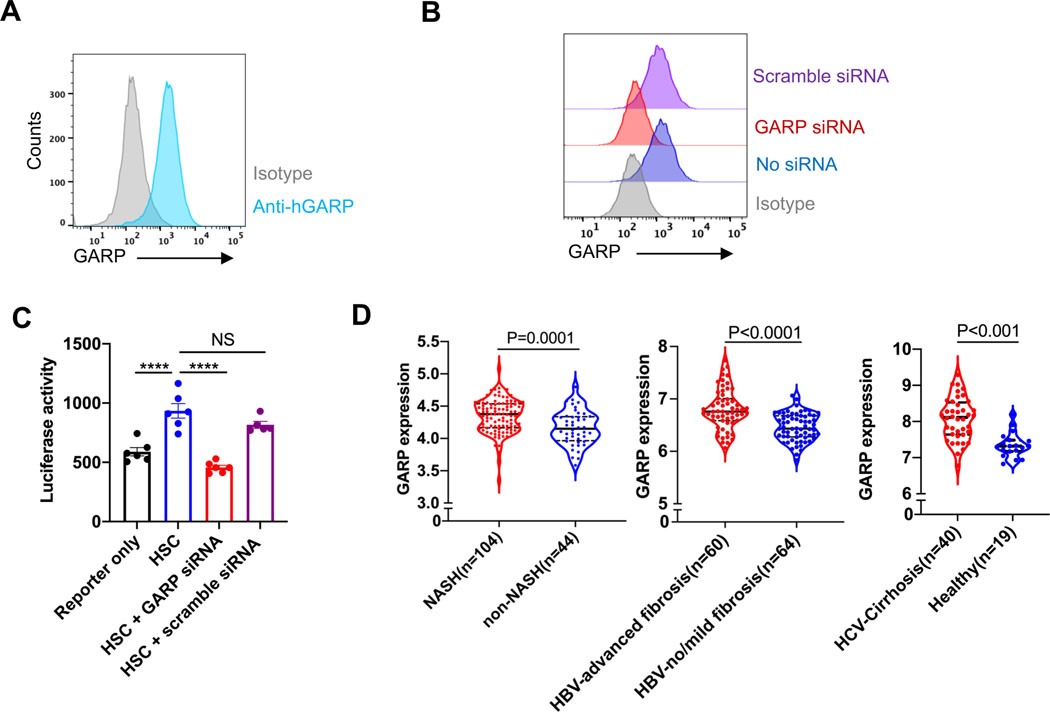
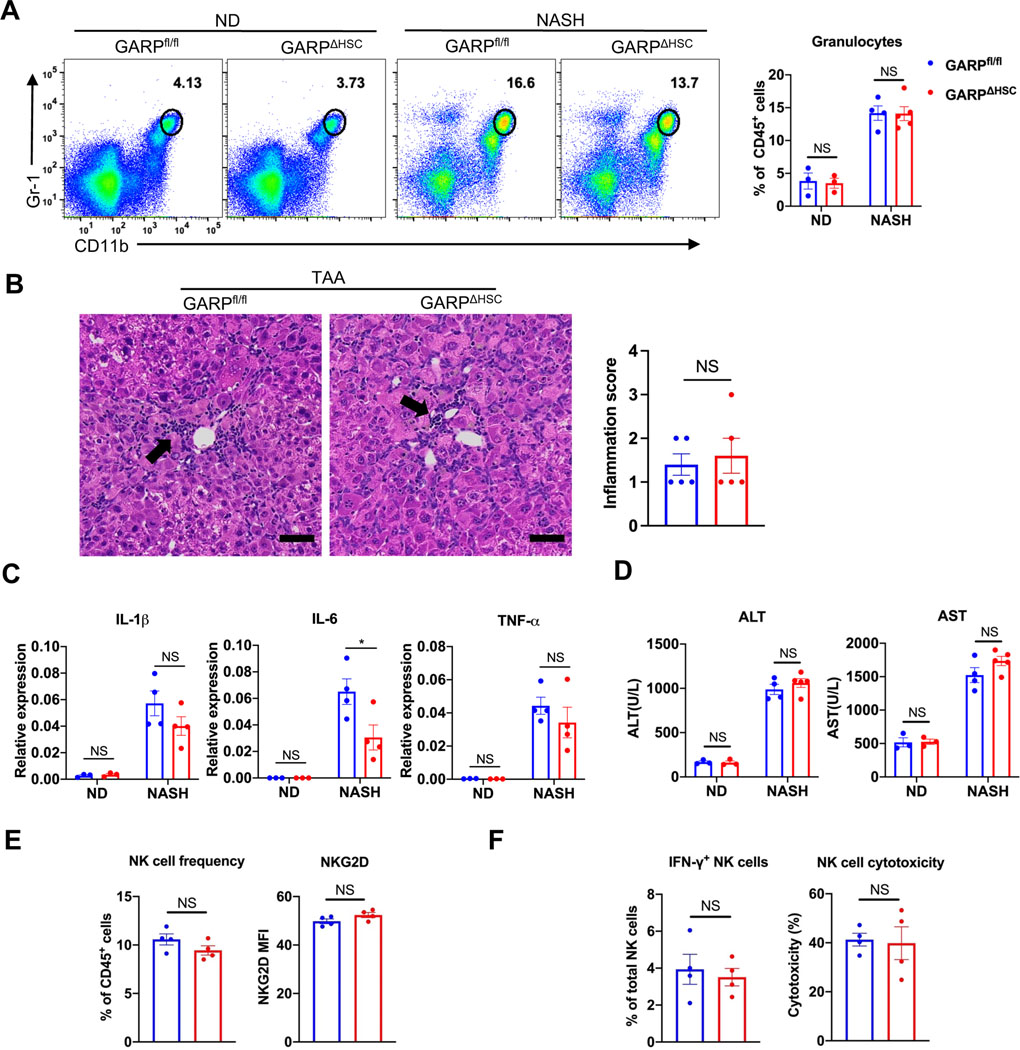
Results: Mice lacking GARP in HSCs were protected from developing liver fibrosis. Therapeutically deleting GARP on HSCs alleviated the fibrotic process in established disease. Furthermore, natural killer T cells exacerbated hepatic fibrosis by inducing GARP expression on HSCs through IL-4 production. Mechanistically, GARP facilitated fibrogenesis by activating TGF-β and enhancing endothelin-1-mediated HSC contraction. Functional GARP was expressed on human HSCs and significantly upregulated in the livers of patients with fibrosis. Lastly, deletion of GARP on HSCs did not augment inflammation or liver damage.
Conclusions: GARP expressed on HSCs drives the development of liver fibrosis via cell contraction-mediated activation of latent TGF-β. Considering that systemic blockade of TGF-β has major side effects, we highlight a therapeutic niche provided by GARP and surface-mediated TGF-β activation. Thus, our findings suggest an important role of GARP on HSCs as a promising target for the treatment of liver fibrosis.
Impact and implications: Liver fibrosis represents a substantial and increasing public health burden globally, for which specific treatments are not available. Glycoprotein A repetitions predominant (GARP) is a membrane protein that functions as a latent TGF-β docking molecule. Here, we show that GARP expressed on hepatic stellate cells drives the development of liver fibrosis. Our findings suggest GARP as a novel target for the treatment of fibrotic disease.
Keywords: GARP; TGF-β; endothelin-1 (ET-1); hepatic stellate cells (HSCs); liver fibrosis; natural killer T (NKT) cells.
Copyright © 2023 European Association for the Study of the Liver. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
-
- Ekstedt M, Hagstrom H, Nasr P, Fredrikson M, Stal P, Kechagias S, et al. Fibrosis stage is the strongest predictor for disease-specific mortality in NAFLD after up to 33 years of follow-up. Hepatology 2015;61:1547–1554. - PubMed
-
- Sepanlou SG, Safiri S, Bisignano C, Ikuta KS, Merat S, Saberifiroozi M, et al. The global, regional, and national burden of cirrhosis by cause in 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. The Lancet Gastroenterology & Hepatology 2020;5:245–266. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
