

Fusarium diversity associated with diseased cereals in China, with an updated phylogenomic assessment of the genus
- PMID: 37351543
- PMCID: PMC10282163
- DOI: 10.3114/sim.2022.104.02
Fusarium diversity associated with diseased cereals in China, with an updated phylogenomic assessment of the genus
Abstract
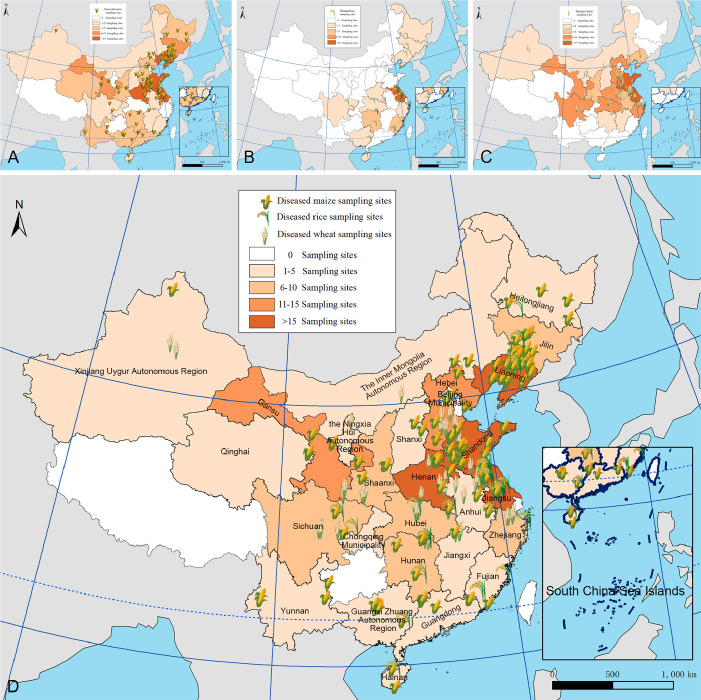
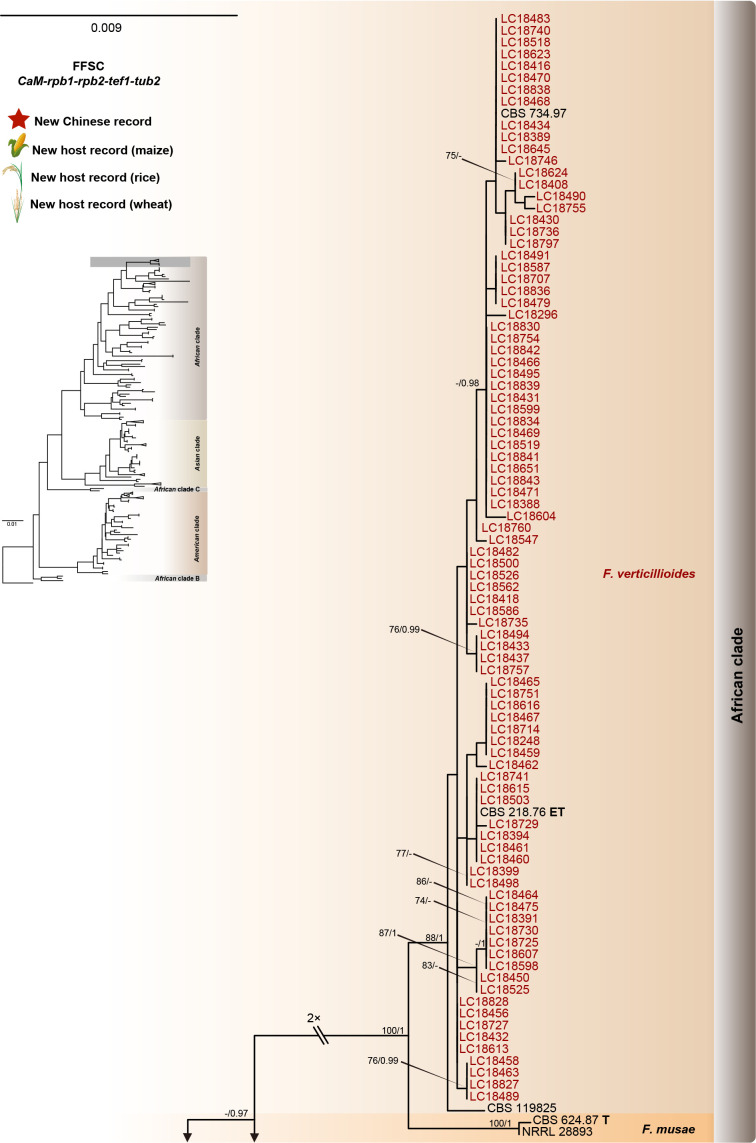
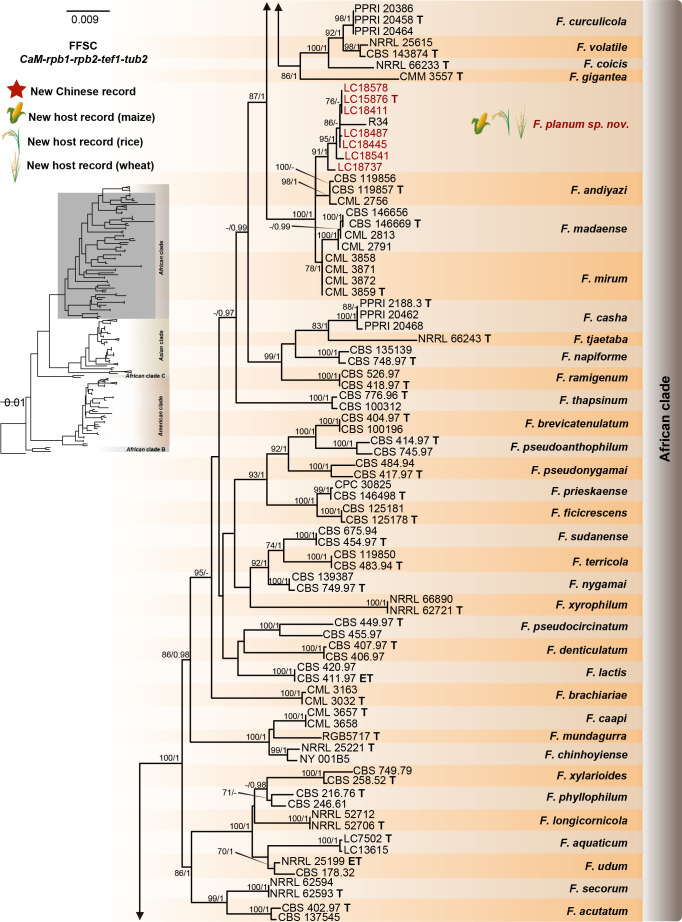
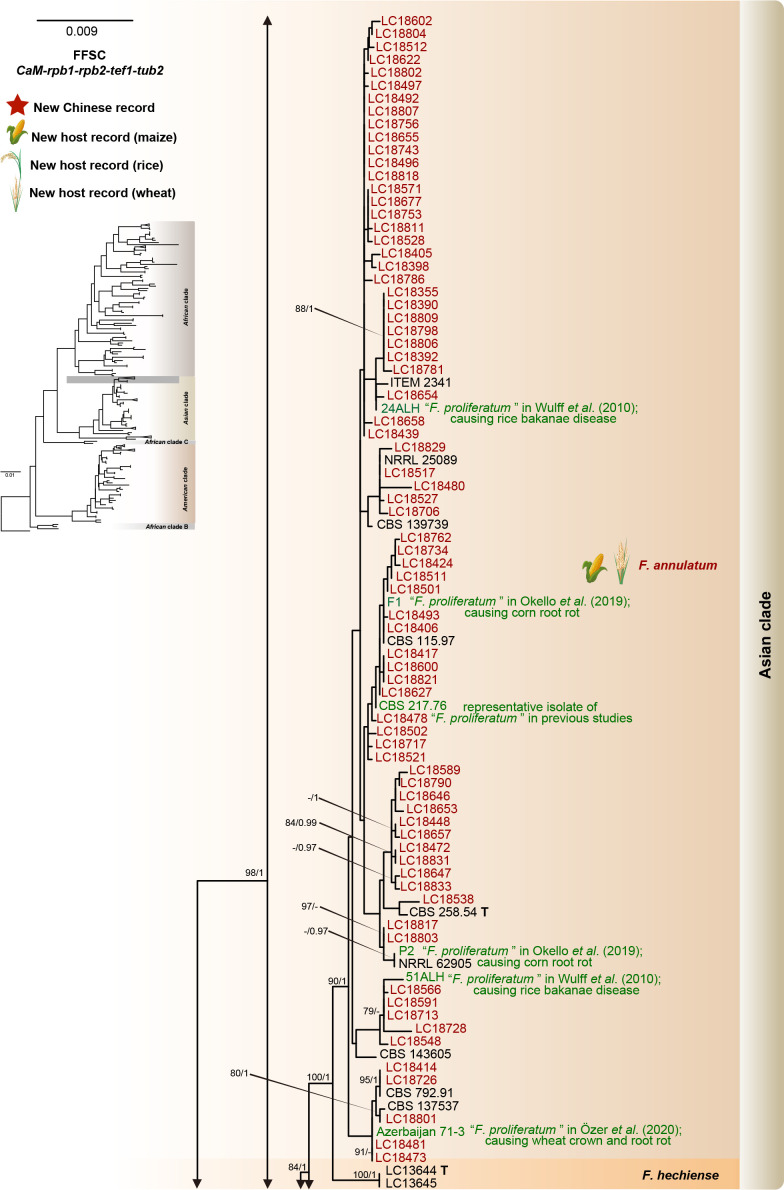
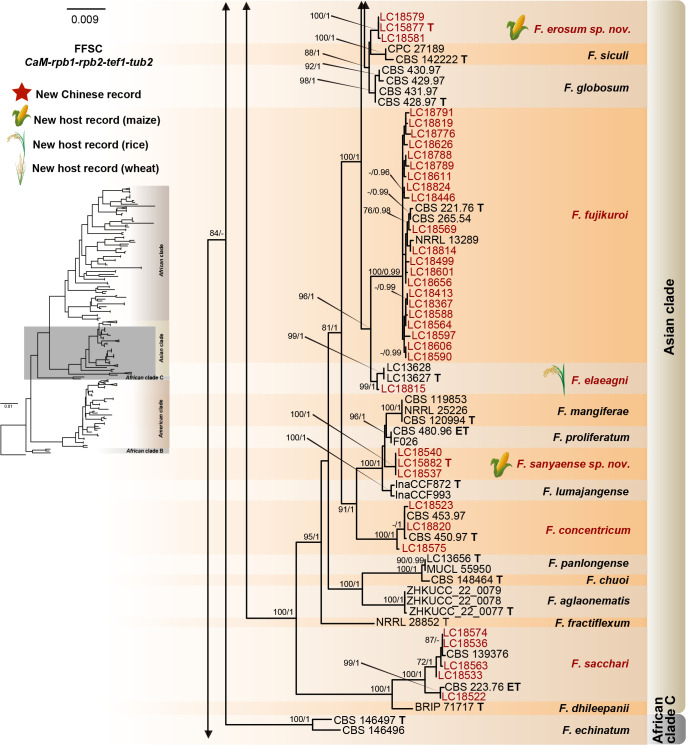
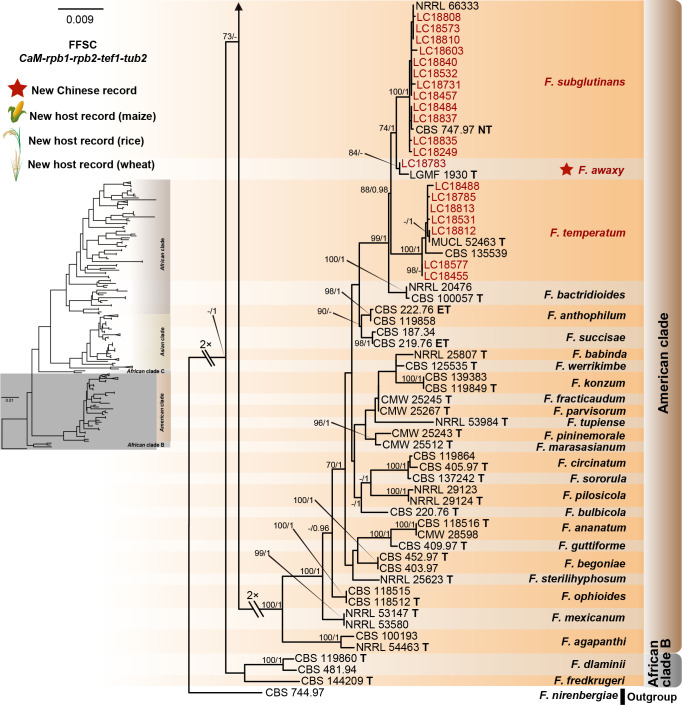
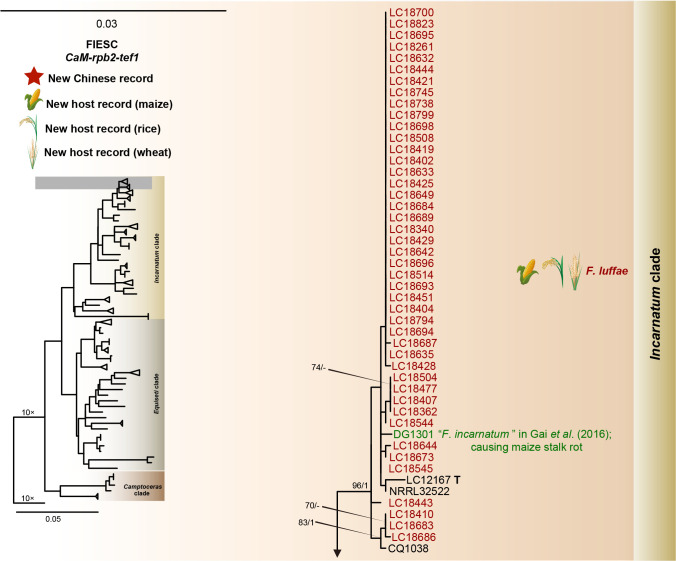
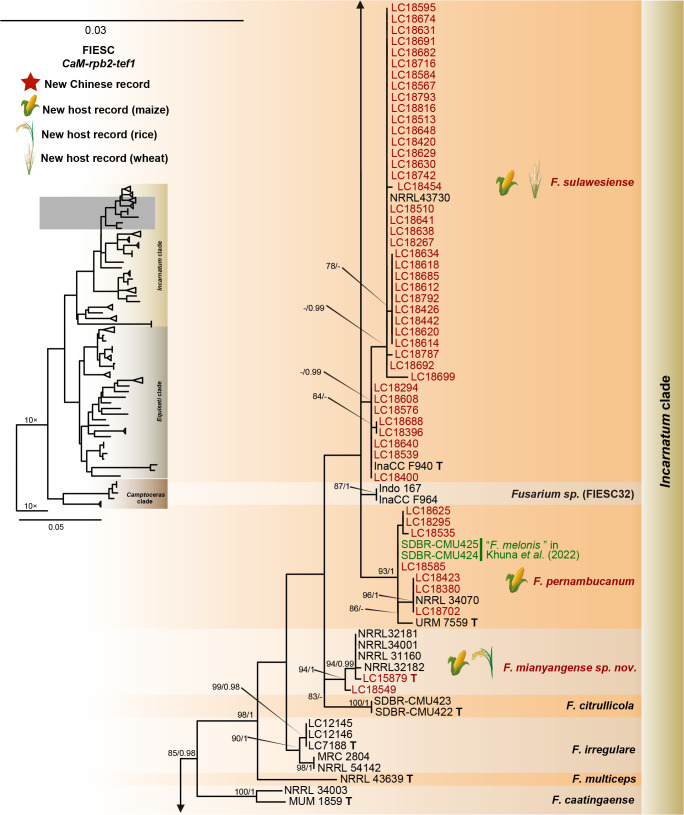
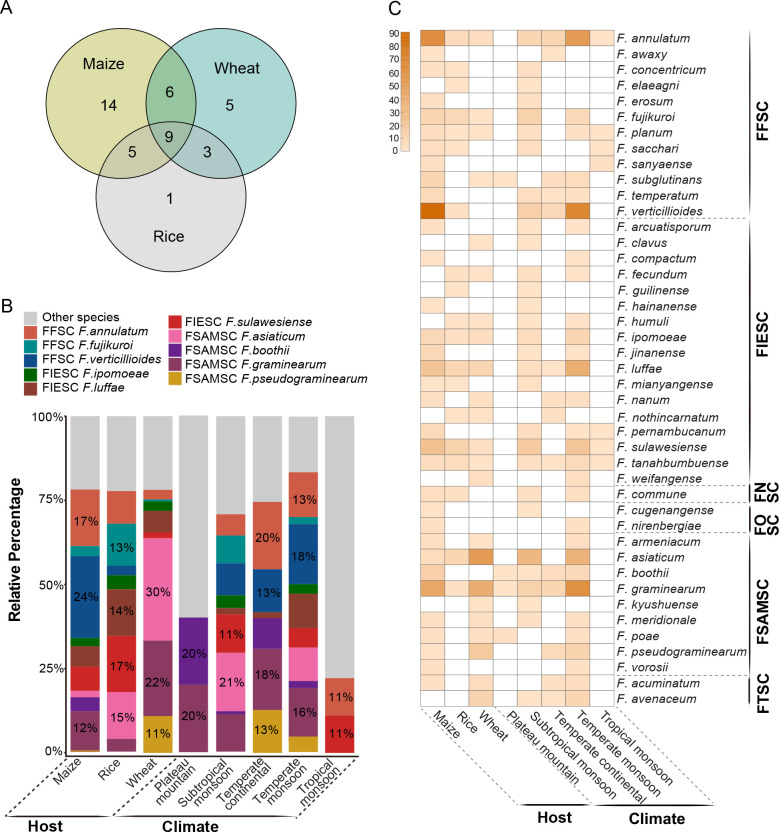
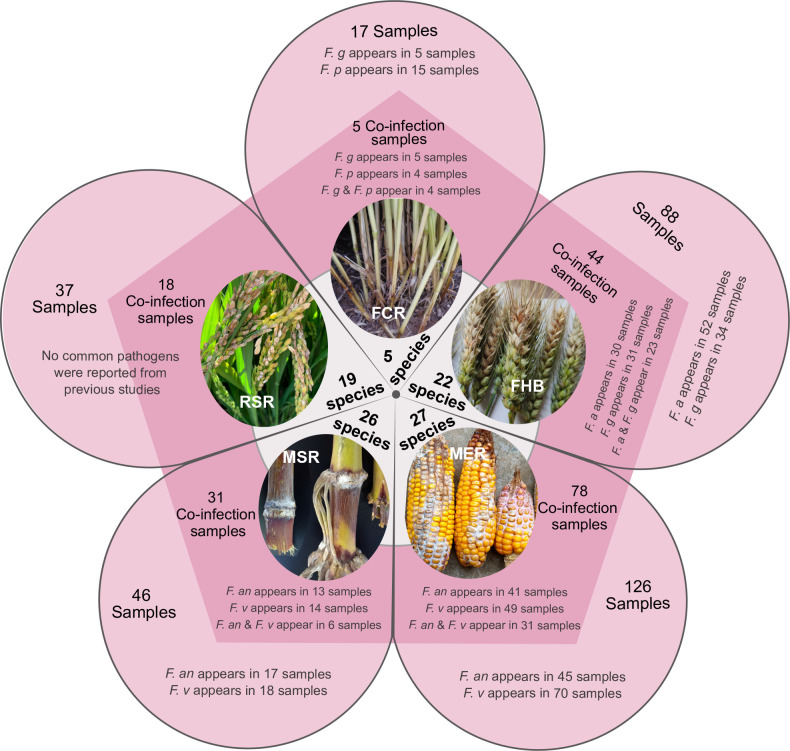
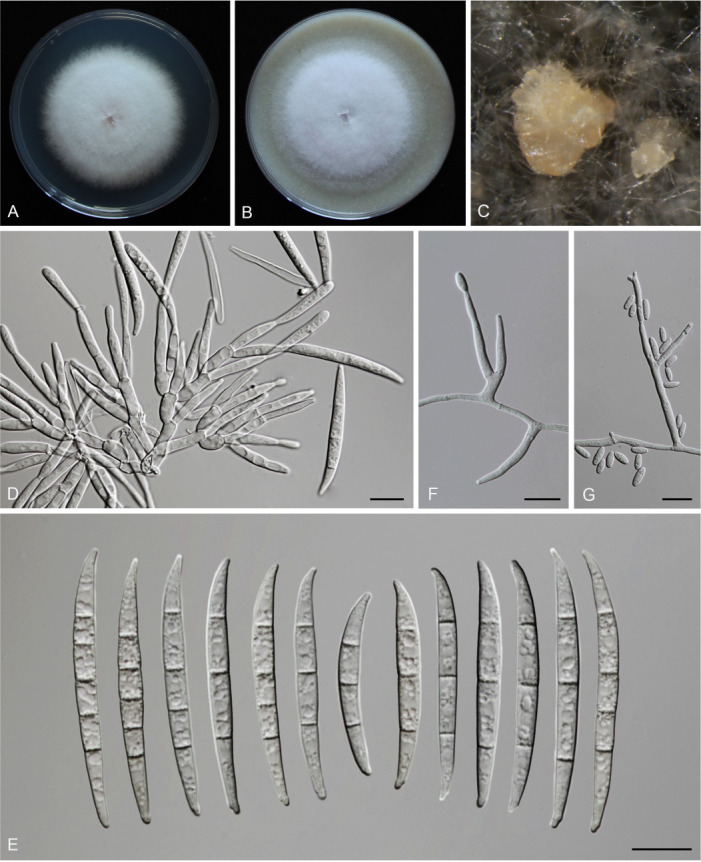
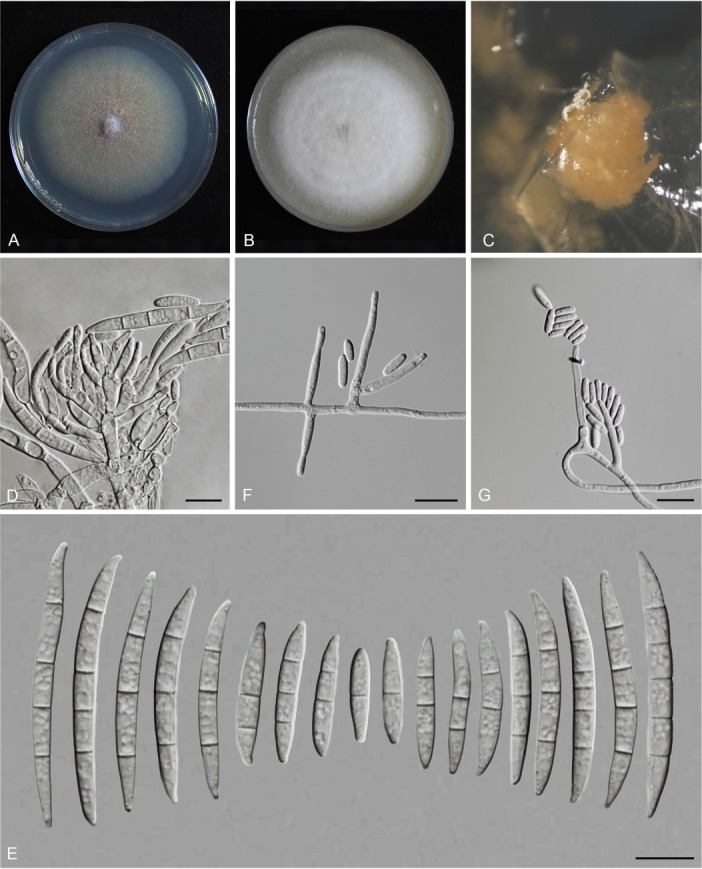
Fusarium species are important cereal pathogens that cause severe production losses to major cereal crops such as maize, rice, and wheat. However, the causal agents of Fusarium diseases on cereals have not been well documented because of the difficulty in species identification and the debates surrounding generic and species concepts. In this study, we used a citizen science initiative to investigate diseased cereal crops (maize, rice, wheat) from 250 locations, covering the major cereal-growing regions in China. A total of 2 020 Fusarium strains were isolated from 315 diseased samples. Employing multi-locus phylogeny and morphological features, the above strains were identified to 43 species, including eight novel species that are described in this paper. A world checklist of cereal-associated Fusarium species is provided, with 39 and 52 new records updated for the world and China, respectively. Notably, 56 % of samples collected in this study were observed to have co-infections of more than one Fusarium species, and the detailed associations are discussed. Following Koch's postulates, 18 species were first confirmed as pathogens of maize stalk rot in this study. Furthermore, a high-confidence species tree was constructed in this study based on 1 001 homologous loci of 228 assembled genomes (40 genomes were sequenced and provided in this study), which supported the "narrow" generic concept of Fusarium (= Gibberella). This study represents one of the most comprehensive surveys of cereal Fusarium diseases to date. It significantly improves our understanding of the global diversity and distribution of cereal-associated Fusarium species, as well as largely clarifies the phylogenetic relationships within the genus. Taxonomic novelties: New species: Fusarium erosum S.L. Han, M.M. Wang & L. Cai, Fusarium fecundum S.L. Han, M.M. Wang & L. Cai, Fusarium jinanense S.L. Han, M.M. Wang & L. Cai, Fusarium mianyangense S.L. Han, M.M. Wang & L. Cai, Fusarium nothincarnatum S.L. Han, M.M. Wang & L. Cai, Fusarium planum S.L. Han, M.M. Wang & L. Cai, Fusarium sanyaense S.L. Han, M.M. Wang & L. Cai, Fusarium weifangense S.L. Han, M.M. Wang & L. Cai. Citation: Han SL, Wang MM, Ma ZY, Raza M, Zhao P, Liang JM, Gao M, Li YJ, Wang JW, Hu DM, Cai L (2023). Fusarium diversity associated with diseased cereals in China, with an updated phylogenomic assessment of the genus. Studies in Mycology 104: 87-148. doi: 10.3114/sim.2022.104.02.
Keywords: Cereal pathogens; citizen science; co-infection; new taxa; pathobiome; phylogeny; species complexes; systematics.
© 2023 Westerdijk Fungal Biodiversity Institute.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures






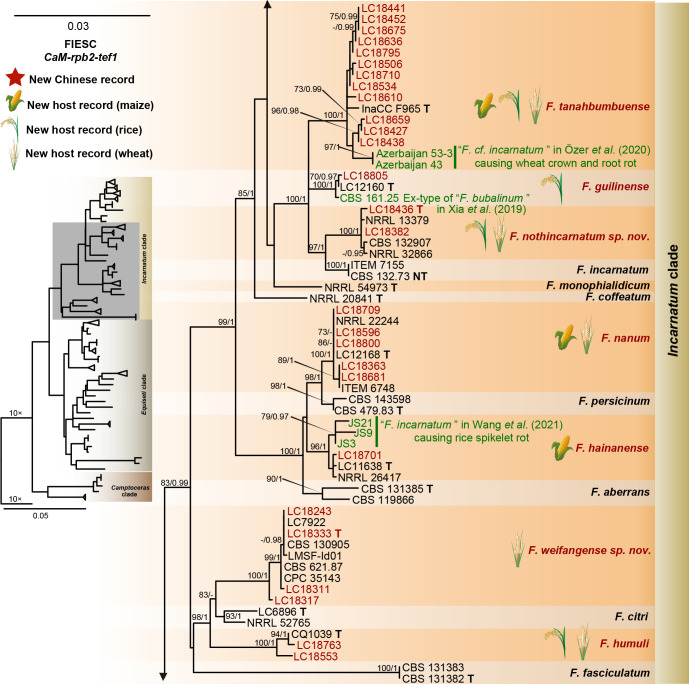
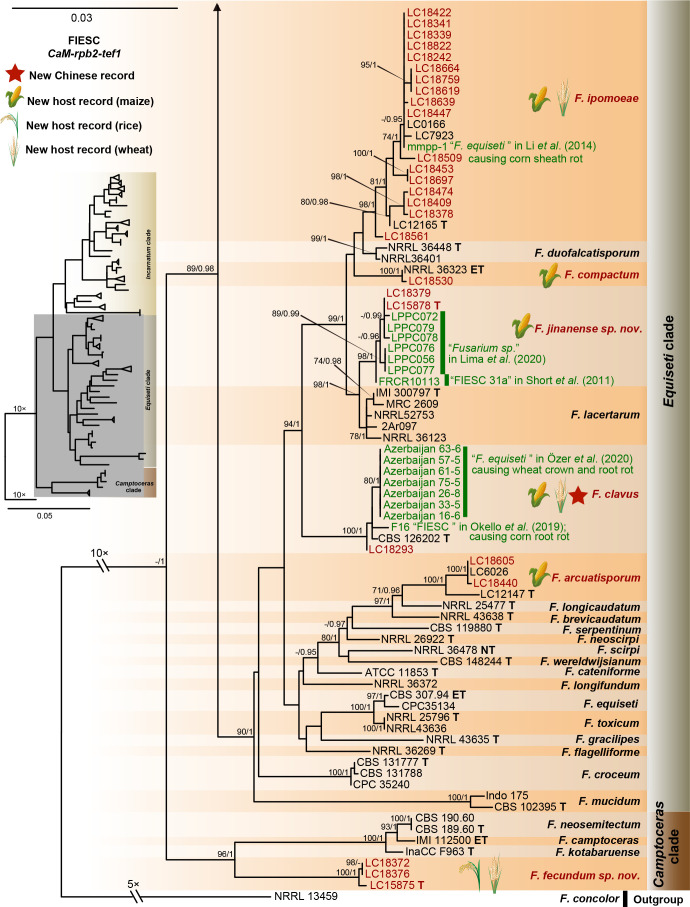
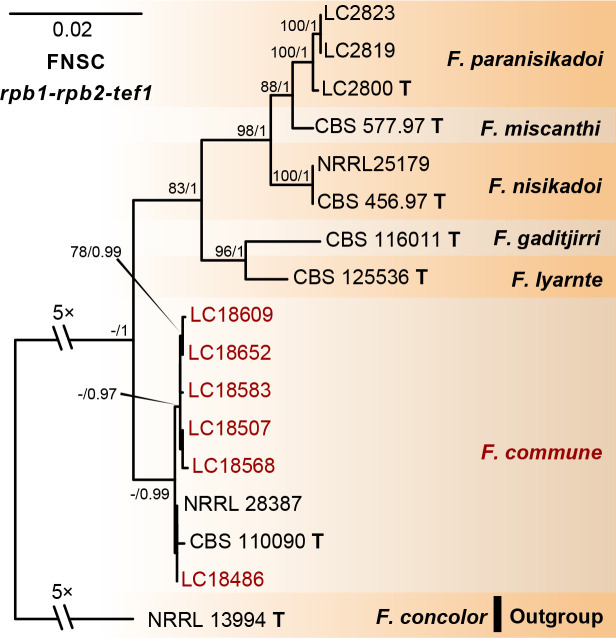
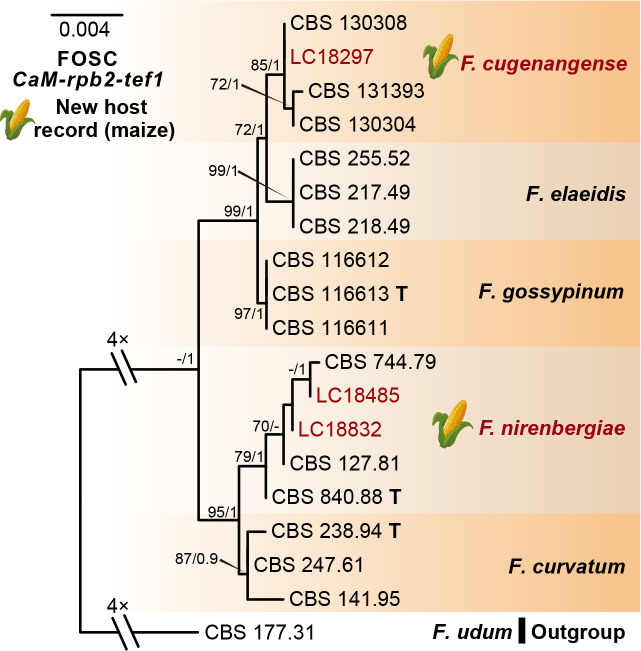
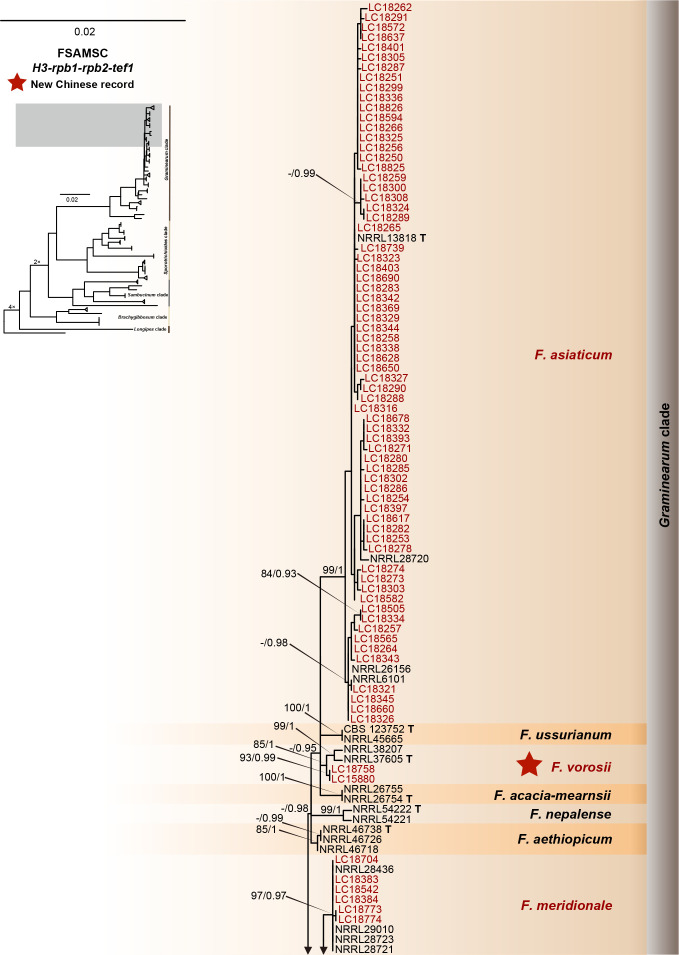
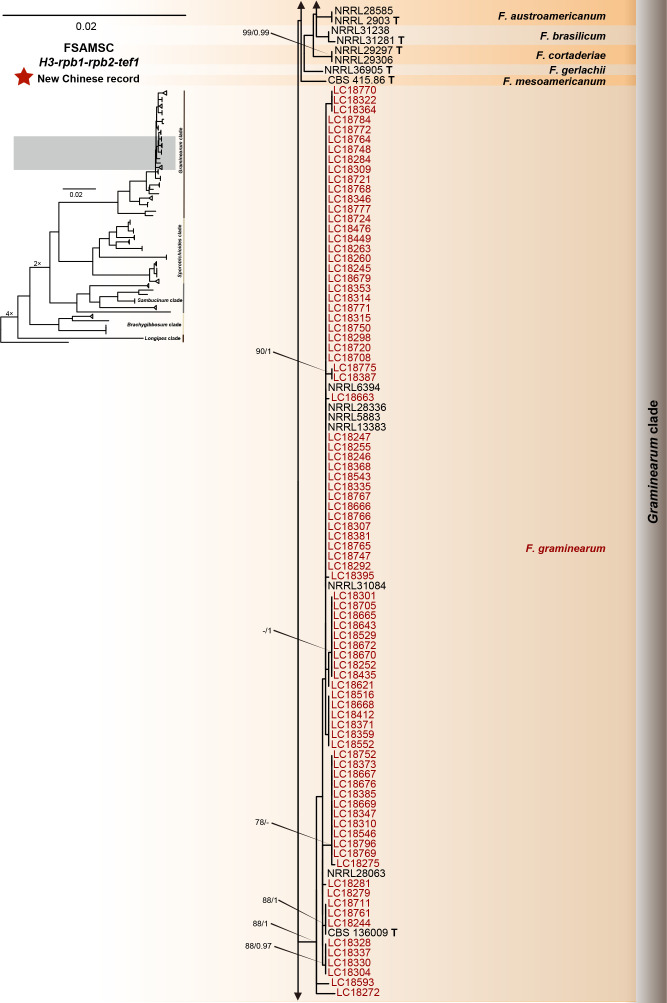
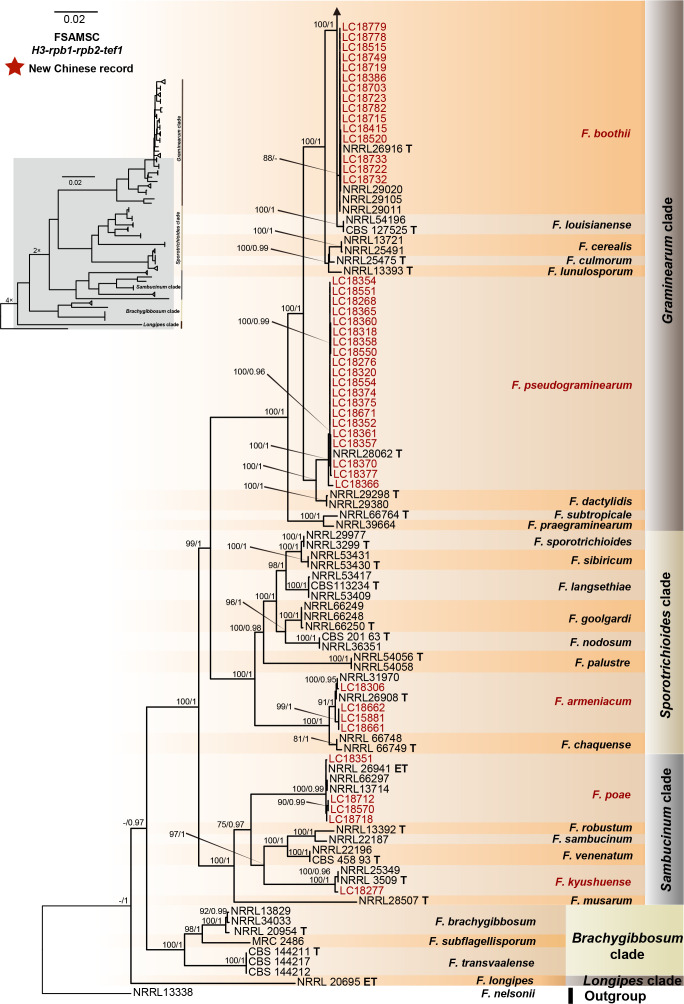
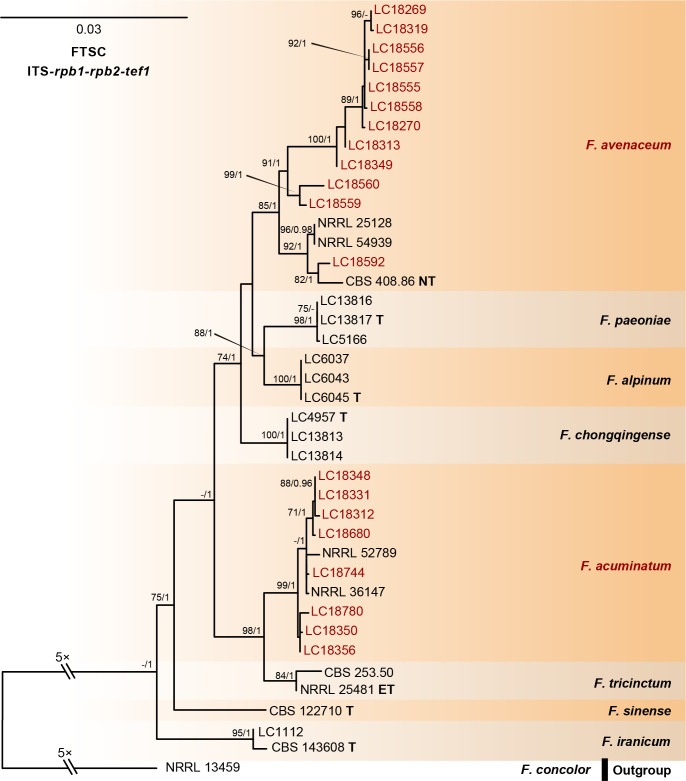
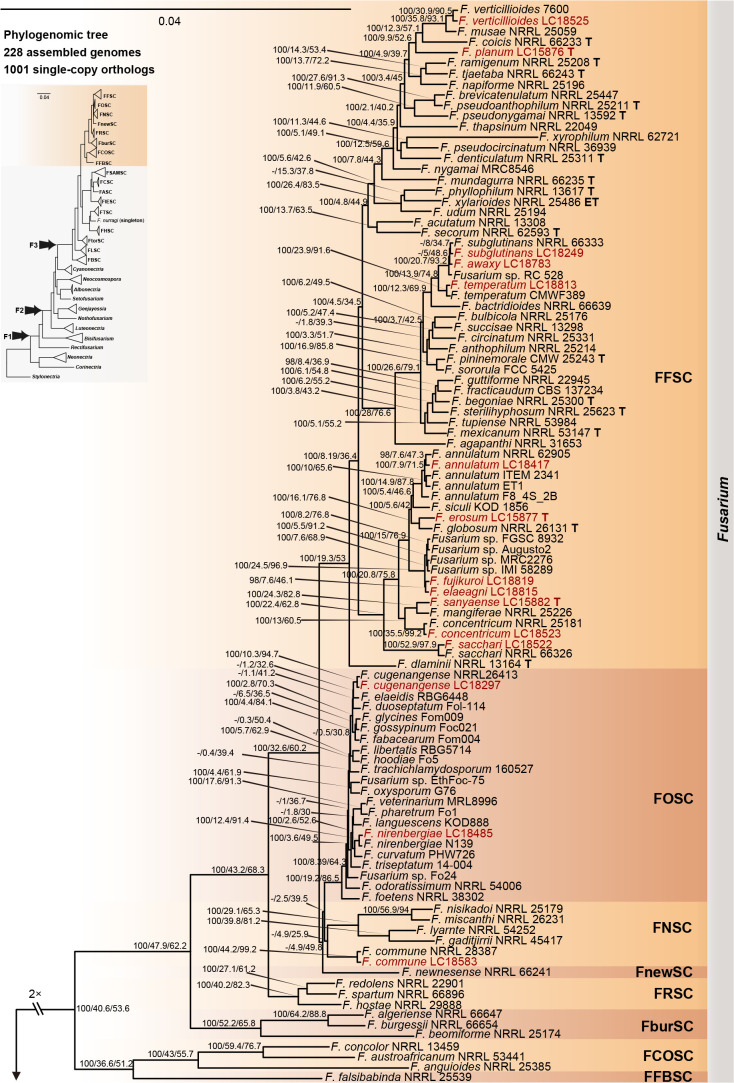
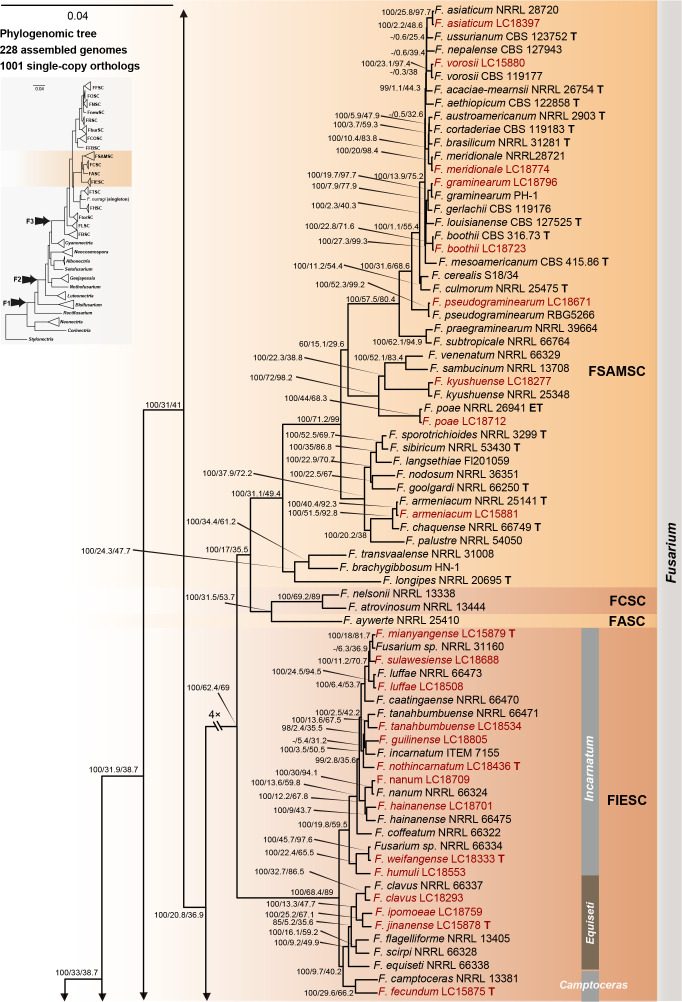
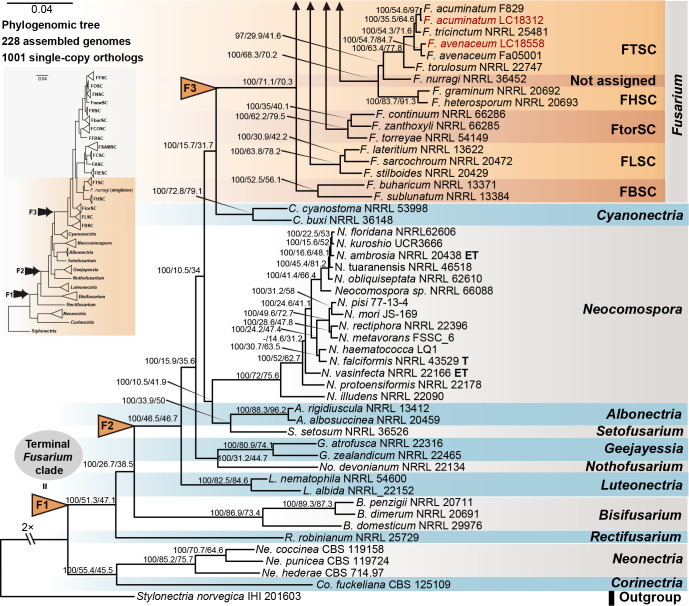
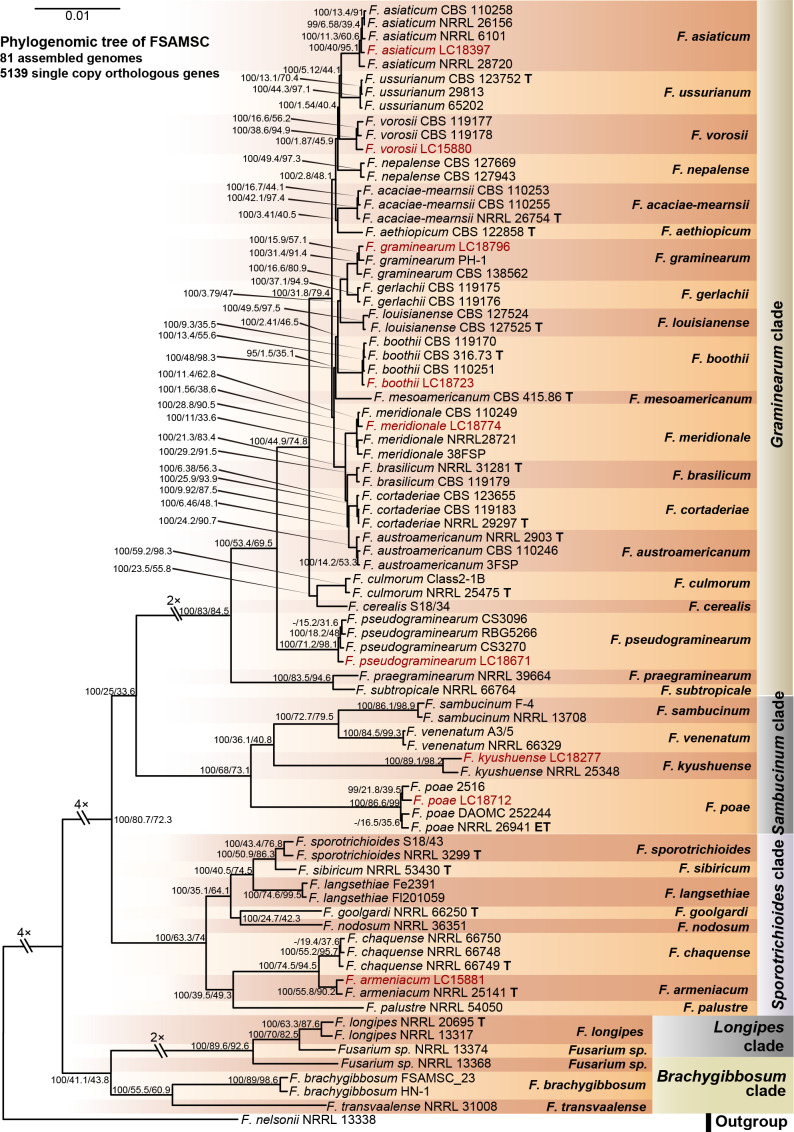
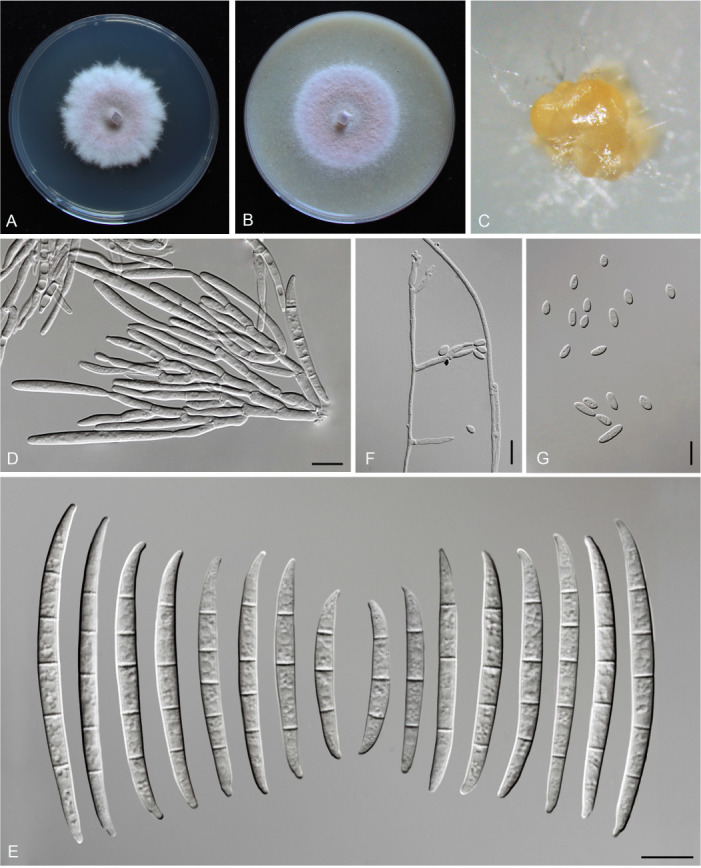
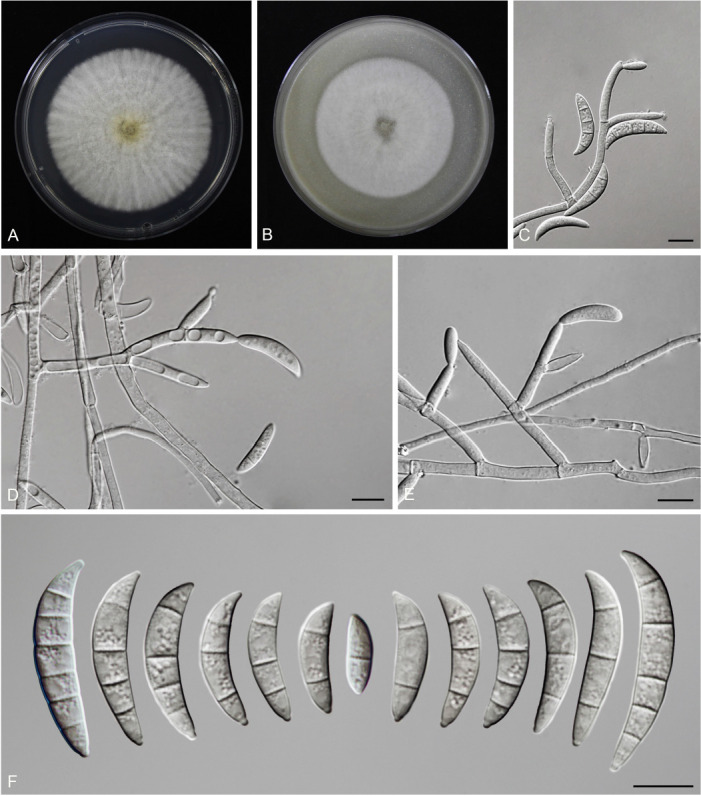
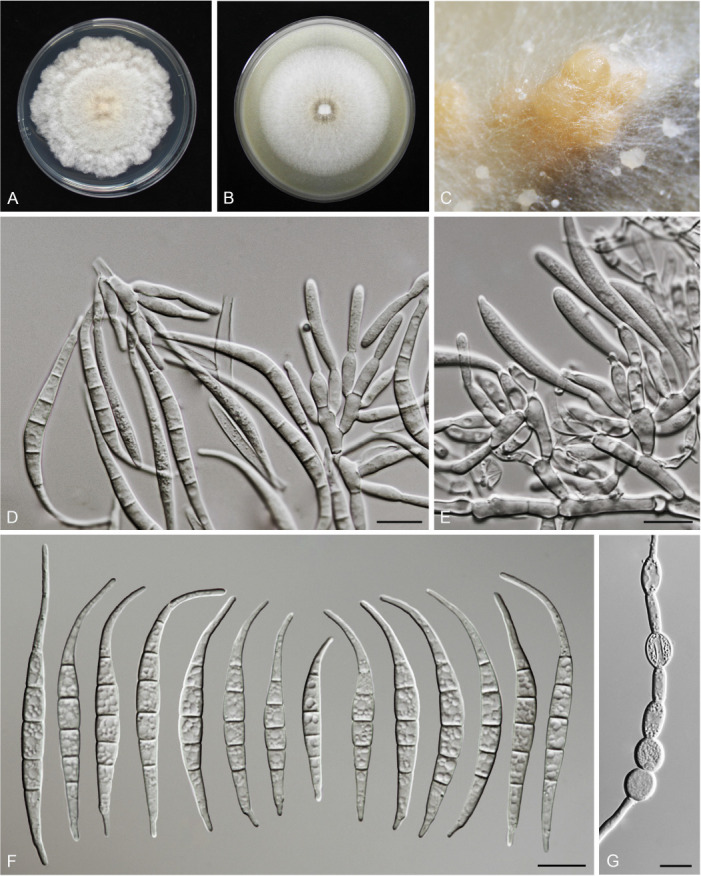
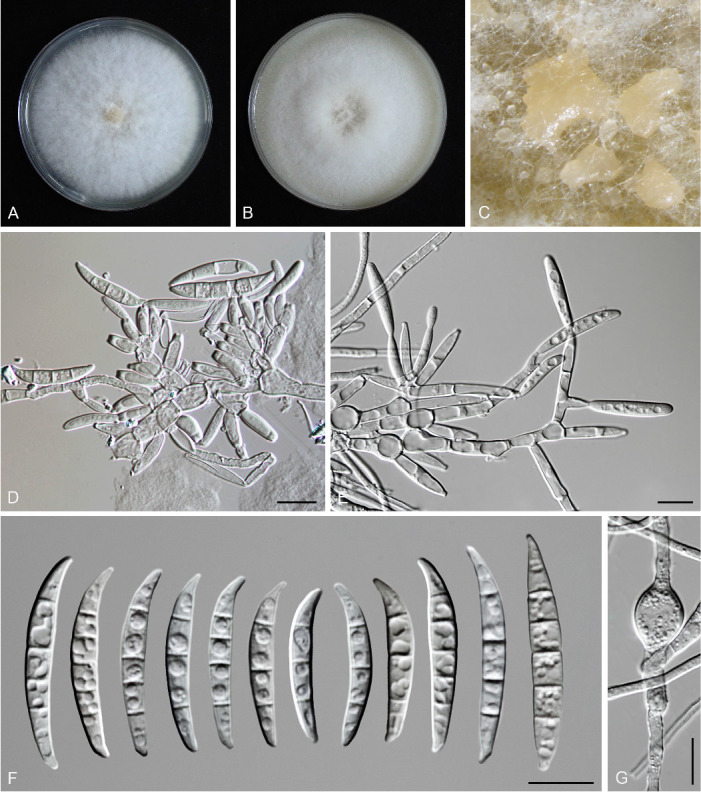
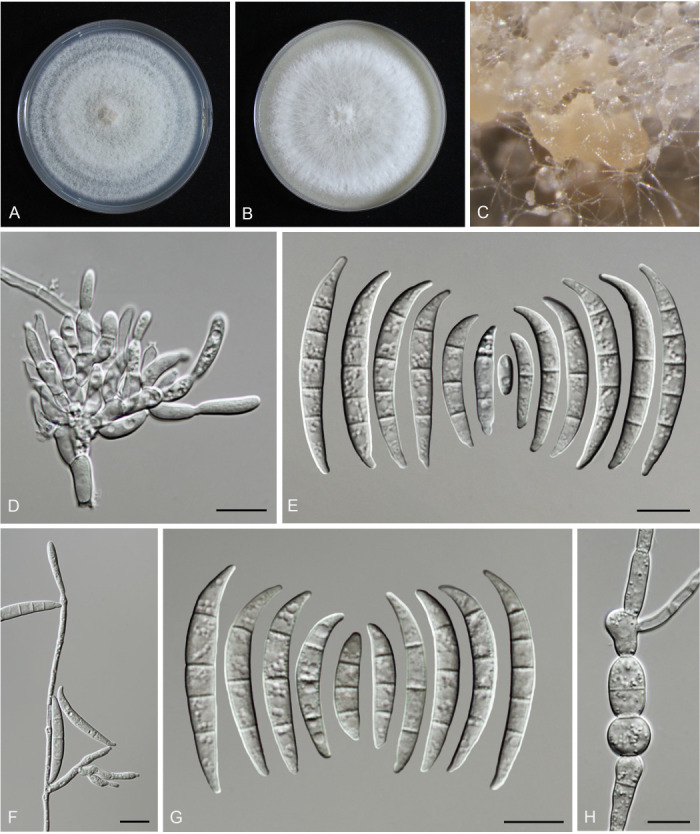
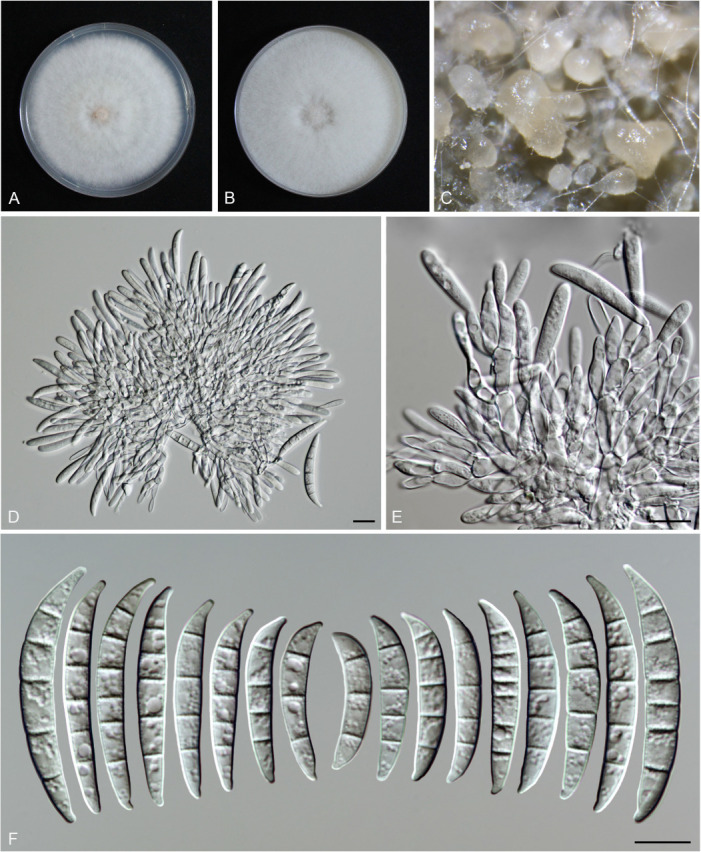
References
-
- Aamot HU, Ward TJ, Brodal G, et al. (2015). Genetic and phenotypic diversity within the Fusarium graminearum species complex in Norway. European Journal of Plant Pathology 142: 501–519.
-
- Abdallah-Nekache N, Laraba I, Ducos C, et al. (2019). Occurrence of Fusarium head blight and Fusarium crown rot in Algerian wheat: identification of associated species and assessment of aggressiveness. European Journal of Plant Pathology 154: 499–512.
-
- Abdul Rahm M, Mohamed Gh K, Essia Reha NA. (2020). Prevalence and transmission of seed-borne fungi of maize and their control by phenolic antioxidants. Plant Pathology Journal 19: 176–184.
-
- Adikaram NKB, Yakandawala DMD. (2020). A checklist of plant pathogenic fungi and Oomycota in Sri Lanka. Ceylon Journal of Science 49: 93–123.
-
- Aguín O, Cao A, Pintos C, et al. (2014). Occurrence of Fusarium species in maize kernels grown in north western Spain. Plant Pathology 63: 946–951.
LinkOut - more resources
Full Text Sources