

Domestication has altered the ABA and gibberellin profiles in developing pea seeds
- PMID: 37351659
- PMCID: PMC10290032
- DOI: 10.1007/s00425-023-04184-2
Domestication has altered the ABA and gibberellin profiles in developing pea seeds
Abstract
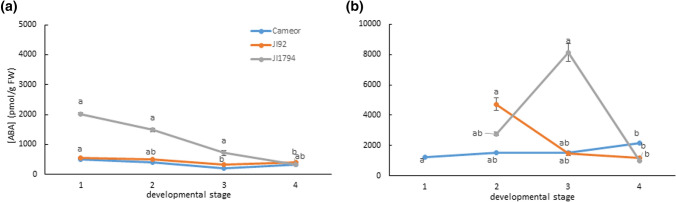
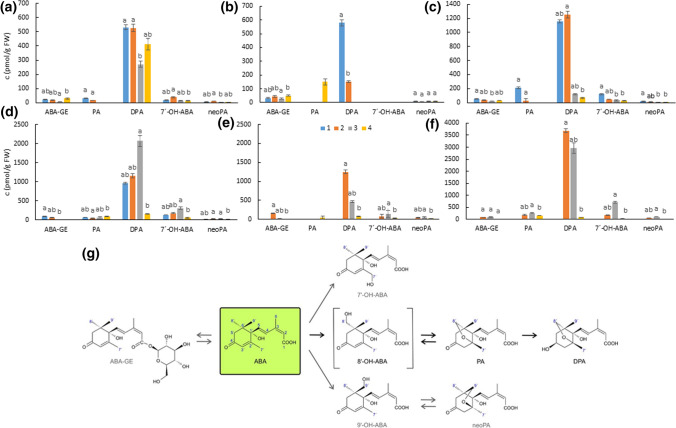
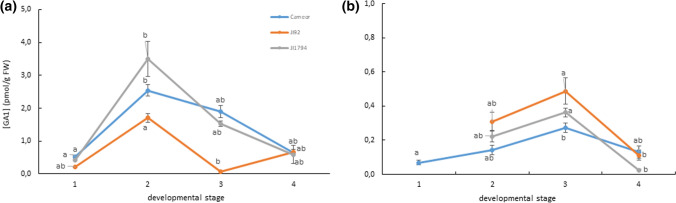
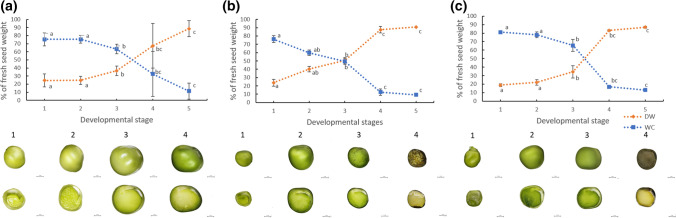
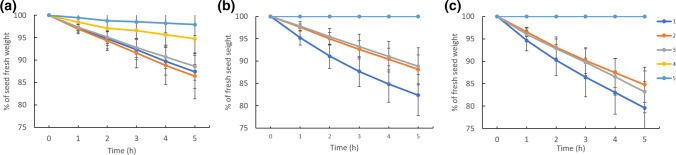
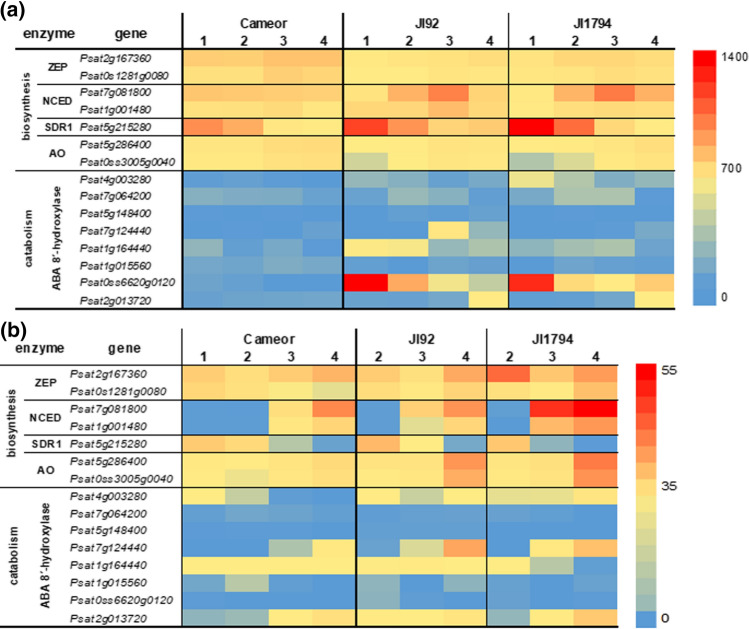
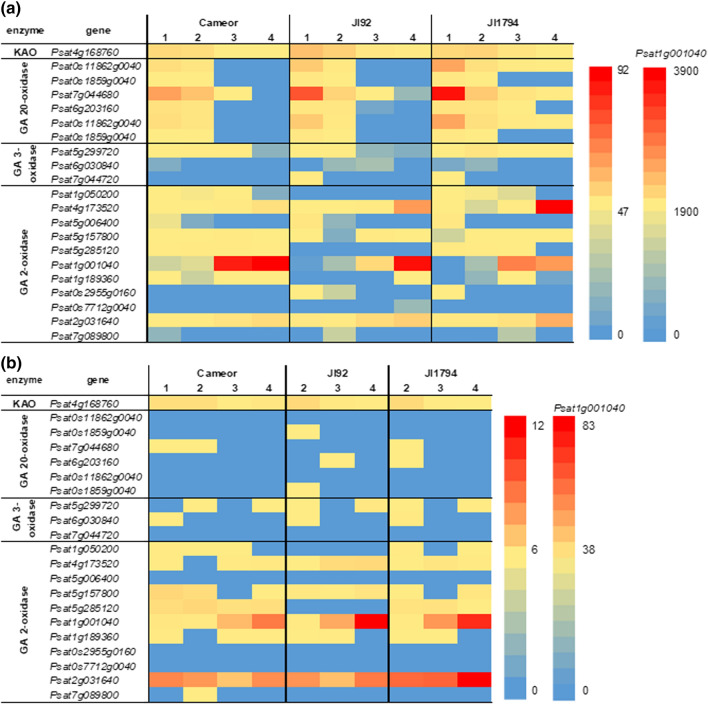
We showed that wild pea seeds contained a more diverse combination of bioactive GAs and had higher ABA content than domesticated peas. Although the role of abscisic acid (ABA) and gibberellins (GAs) interplay has been extensively studied in Arabidopsis and cereals models, comparatively little is known about the effect of domestication on the level of phytohormones in legume seeds. In legumes, as in other crops, seed dormancy has been largely or entirely removed during domestication. In this study, we have measured the endogenous levels of ABA and GAs comparatively between wild and domesticated pea seeds during their development. We have shown that wild seeds contained more ABA than domesticated ones, which could be important for preparing the seeds for the period of dormancy. ABA was catabolised particularly by an 8´-hydroxylation pathway, and dihydrophaseic acid was the main catabolite in seed coats as well as embryos. Besides, the seed coats of wild and pigmented cultivated genotypes were characterised by a broader spectrum of bioactive GAs compared to non-pigmented domesticated seeds. GAs in both seed coat and embryo were synthesized mainly by a 13-hydroxylation pathway, with GA29 being the most abundant in the seed coat and GA20 in the embryos. Measuring seed water content and water loss indicated domesticated pea seeds´ desiccation was slower than that of wild pea seeds. Altogether, we showed that pea domestication led to a change in bioactive GA composition and a lower ABA content during seed development.
Keywords: Desiccation; Legume; Maturation; Phytohormones; Pigmentation; Seed-coat.
© 2023. The Author(s).
Conflict of interest statement
The authors have no competing interests to declare that are relevant to the content of this article.
Figures




References
-
- Balarynová J, Klčová B, Sekaninová J, Kobrlová L, Cechová MZ, Krejčí P, Leonova T, Gorbach D, Ihling C, Smržová L, Trněný O, Frolov A, Bednář P, Smýkal P. The loss of polyphenol oxidase function is associated with hilum pigmentation and has been selected during pea domestication. New Phytol. 2022;235:1807–1821. doi: 10.1111/nph.18256. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
