

Ablation of microglia following infection of the central nervous system with a neurotropic murine coronavirus infection leads to increased demyelination and impaired remyelination
- PMID: 37352687
- PMCID: PMC11840753
- DOI: 10.1016/j.jneuroim.2023.578133
Ablation of microglia following infection of the central nervous system with a neurotropic murine coronavirus infection leads to increased demyelination and impaired remyelination
Abstract
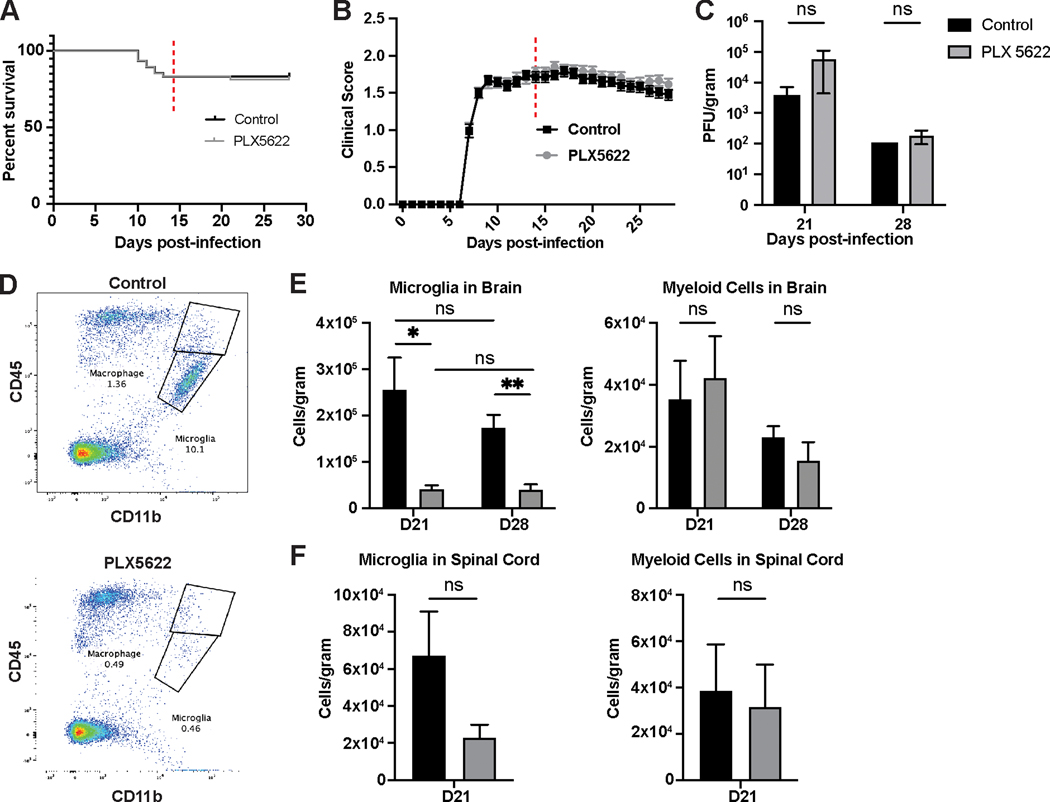
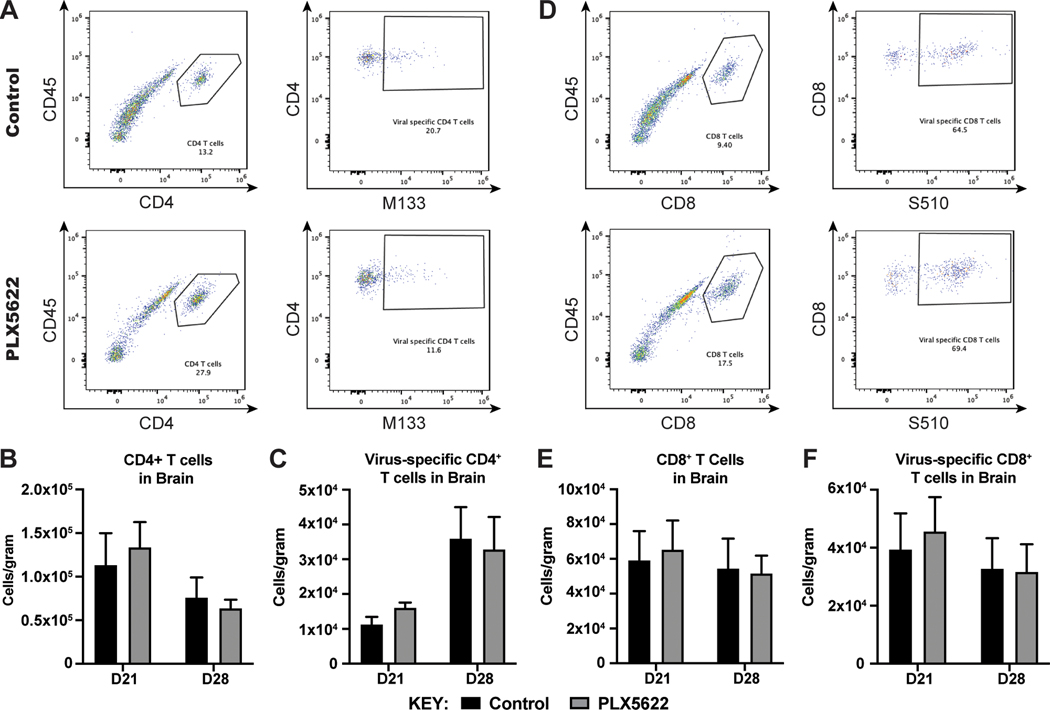
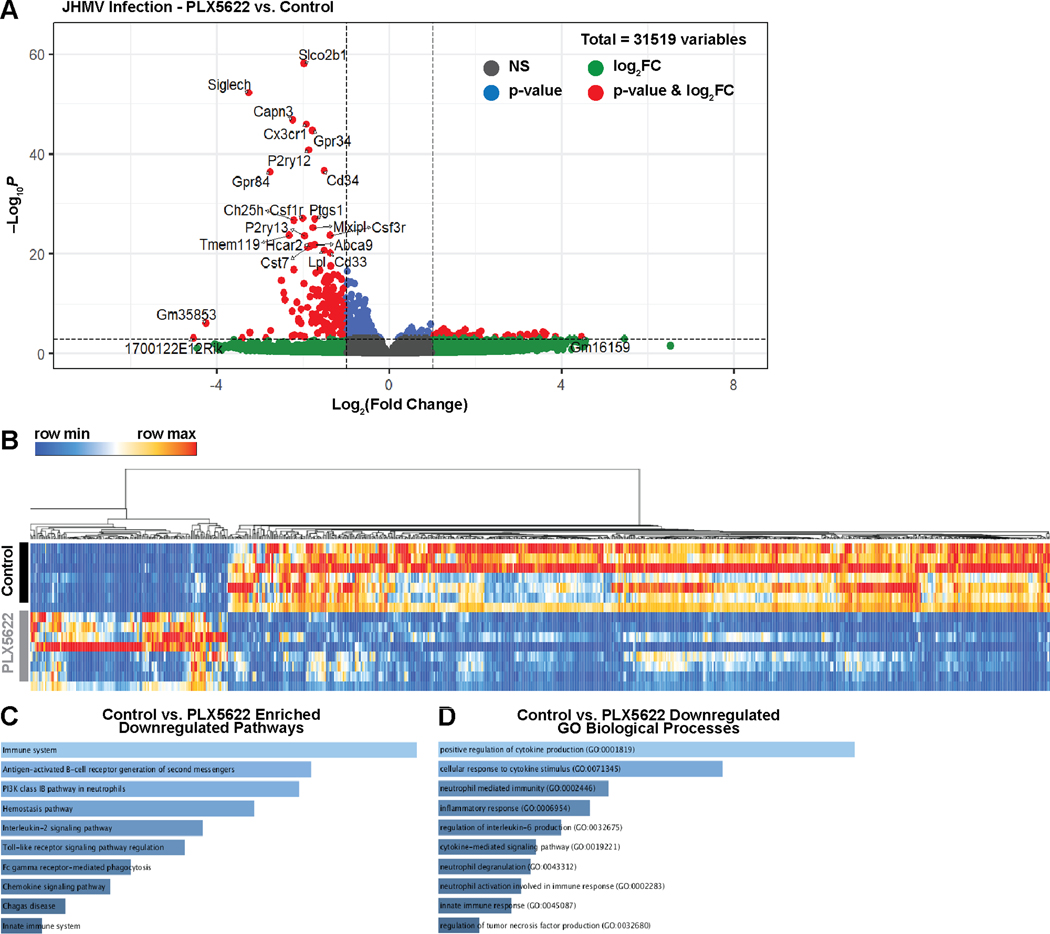
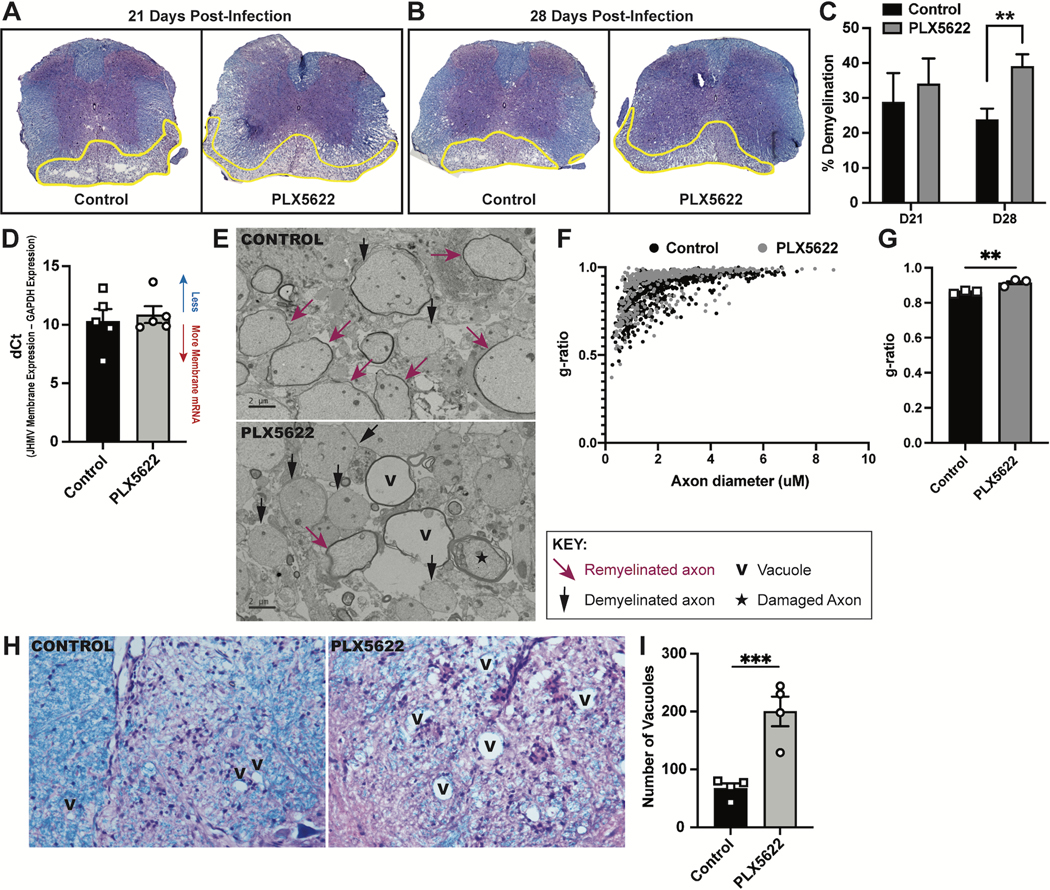
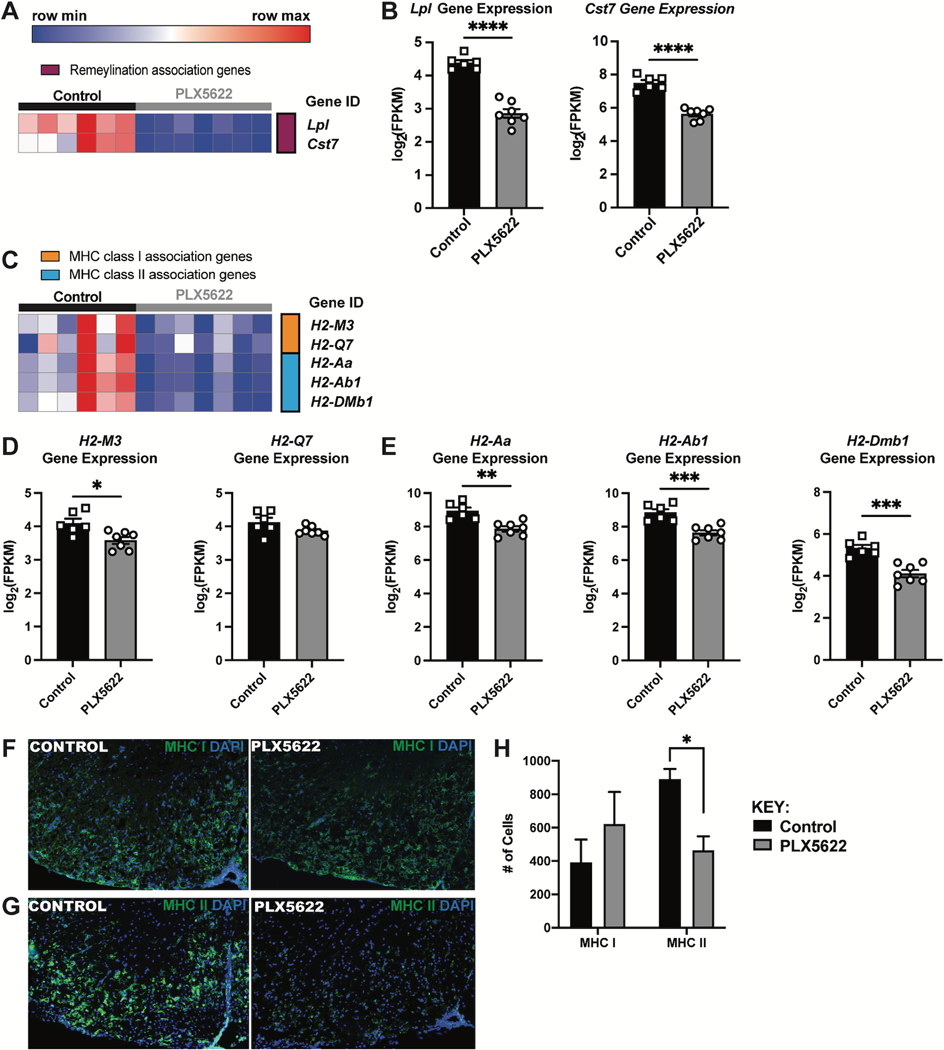
Intracranial inoculation of susceptible mice with a glial-tropic strain of mouse hepatitis virus (JHMV), a murine coronavirus, results in an acute encephalomyelitis followed by viral persistence in white matter tracts accompanied by chronic neuroinflammation and demyelination. Microglia are the resident immune cell of the central nervous system (CNS) and are considered important in regulating events associated with neuroinflammation as well as influencing both white matter damage and remyelination. To better understand mechanisms by which microglia contribute to these immune-mediated events, JHMV-infected mice with established demyelination were treated with the small molecular inhibitor of colony stimulating factor 1 receptor (CSF1R), PLX5622, to deplete microglia. Treatment with PLX5622 did not affect viral replication within the CNS yet the severity of demyelination was increased and remyelination impaired compared to control mice. Gene expression analysis revealed that targeting microglia resulted in altered expression of genes associated with immune cell activation and phagocytosis of myelin debris. These findings indicate that microglia are not critical in viral surveillance in persistently JHMV-infected mice yet restrict white matter damage and remyelination, in part, by influencing phagocytosis of myelin debris.
Keywords: Coronavirus; Demyelination; Microglia; Neuroinflammation; Remyelination.
Copyright © 2023. Published by Elsevier B.V.
Conflict of interest statement
Declaration of Competing Interest None.
Figures
Similar articles
-
Cystatin F attenuates neuroinflammation and demyelination following murine coronavirus infection of the central nervous system.J Neuroinflammation. 2024 Jun 15;21(1):157. doi: 10.1186/s12974-024-03153-0. J Neuroinflammation. 2024. PMID: 38879499 Free PMC article.
-
Microglia influence immune responses and restrict neurologic disease in response to central nervous system infection by a neurotropic murine coronavirus.Front Cell Neurosci. 2023 Nov 30;17:1291255. doi: 10.3389/fncel.2023.1291255. eCollection 2023. Front Cell Neurosci. 2023. PMID: 38099152 Free PMC article. Review.
-
Microglia depletion exacerbates demyelination and impairs remyelination in a neurotropic coronavirus infection.Proc Natl Acad Sci U S A. 2020 Sep 29;117(39):24464-24474. doi: 10.1073/pnas.2007814117. Epub 2020 Sep 14. Proc Natl Acad Sci U S A. 2020. PMID: 32929007 Free PMC article.
-
Microglia influence host defense, disease, and repair following murine coronavirus infection of the central nervous system.Glia. 2020 Nov;68(11):2345-2360. doi: 10.1002/glia.23844. Epub 2020 May 25. Glia. 2020. PMID: 32449994 Free PMC article.
-
The roles of microglia and astrocytes in phagocytosis and myelination: Insights from the cuprizone model of multiple sclerosis.Glia. 2022 Jul;70(7):1215-1250. doi: 10.1002/glia.24148. Epub 2022 Feb 2. Glia. 2022. PMID: 35107839 Free PMC article. Review.
Cited by
-
Cystatin F attenuates neuroinflammation and demyelination following murine coronavirus infection of the central nervous system.J Neuroinflammation. 2024 Jun 15;21(1):157. doi: 10.1186/s12974-024-03153-0. J Neuroinflammation. 2024. PMID: 38879499 Free PMC article.
-
The Contribution of Microglia and Brain-Infiltrating Macrophages to the Pathogenesis of Neuroinflammatory and Neurodegenerative Diseases during TMEV Infection of the Central Nervous System.Viruses. 2024 Jan 13;16(1):119. doi: 10.3390/v16010119. Viruses. 2024. PMID: 38257819 Free PMC article. Review.
-
Microglia influence immune responses and restrict neurologic disease in response to central nervous system infection by a neurotropic murine coronavirus.Front Cell Neurosci. 2023 Nov 30;17:1291255. doi: 10.3389/fncel.2023.1291255. eCollection 2023. Front Cell Neurosci. 2023. PMID: 38099152 Free PMC article. Review.
References
-
- Liu MT, Armstrong D, Hamilton TA, Lane TE. 2001. Expression of Mig (Monokine Induced by Interferon- ) Is Important in T Lymphocyte Recruitment and Host Defense Following Viral Infection of the Central Nervous System. The Journal of Immunology 166:1790–1795. - PubMed
-
- Liu MT, Chen BP, Oertel P, Buchmeier MJ, Armstrong D, Hamilton TA, Lane TE. 2000. Cutting Edge: The T Cell Chemoattractant IFN-Inducible Protein 10 Is Essential in Host Defense Against Viral-Induced Neurologic Disease. The Journal of Immunology 165:2327–2330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
