

Mechanistic insights into the aggregation pathway of the patient-derived immunoglobulin light chain variable domain protein FOR005
- PMID: 37353525
- PMCID: PMC10290123
- DOI: 10.1038/s41467-023-39280-0
Mechanistic insights into the aggregation pathway of the patient-derived immunoglobulin light chain variable domain protein FOR005
Abstract
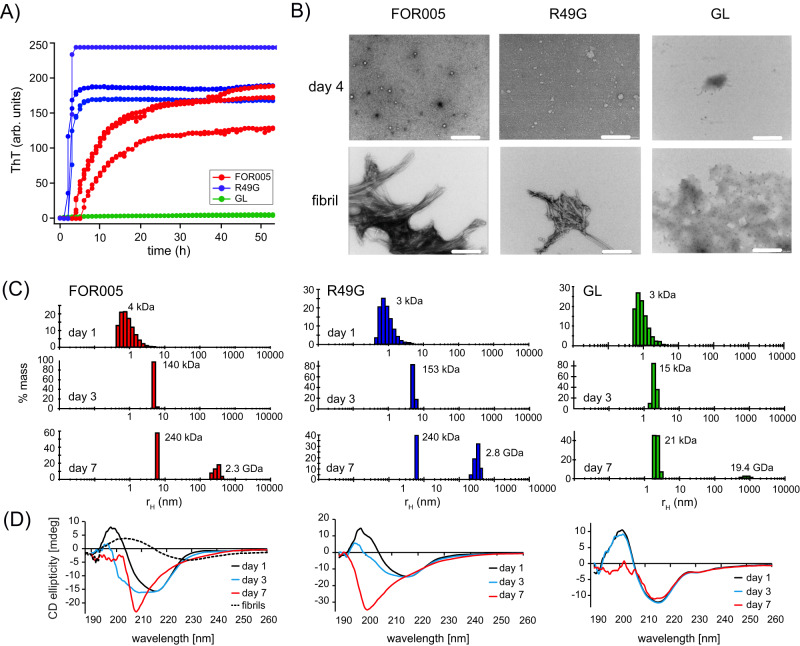
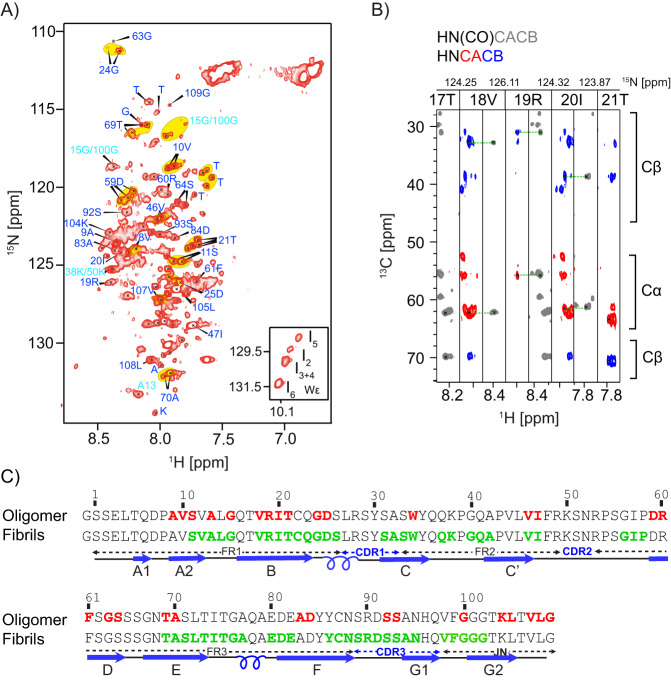
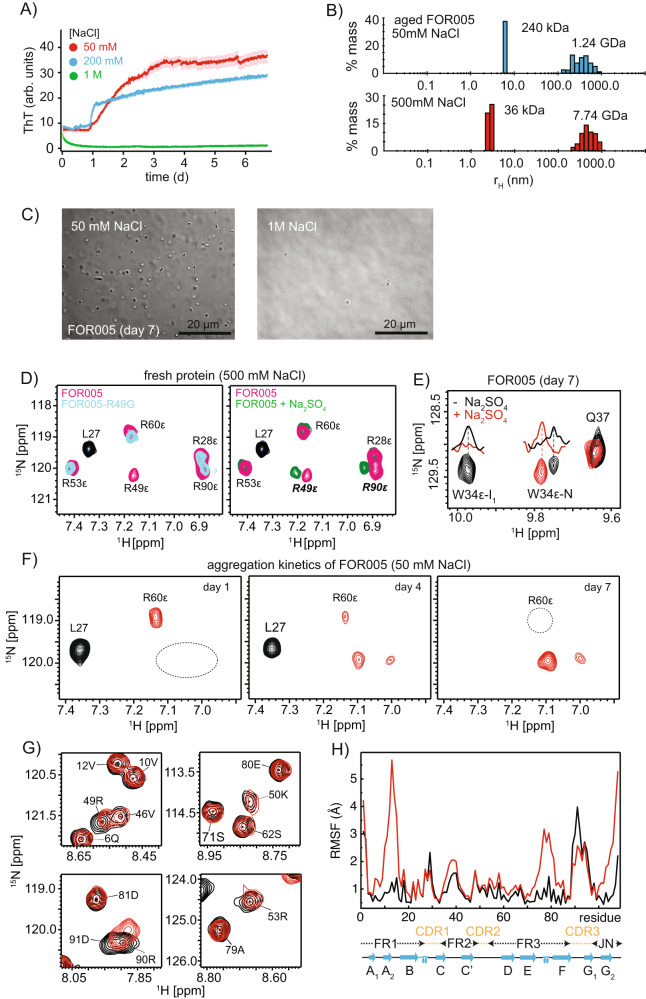
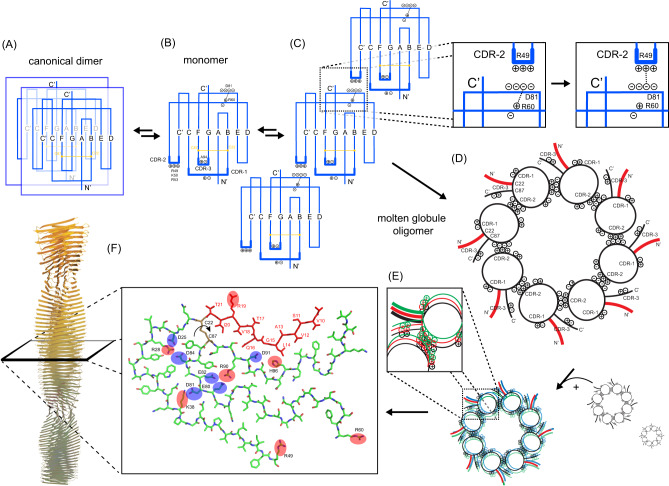
Systemic antibody light chain (AL) amyloidosis is characterized by deposition of amyloid fibrils. Prior to fibril formation, soluble oligomeric AL protein has a direct cytotoxic effect on cardiomyocytes. We focus on the patient derived λ-III AL variable domain FOR005 which is mutated at five positions with respect to the closest germline protein. Using solution-state NMR spectroscopy, we follow the individual steps involved in protein misfolding from the native to the amyloid fibril state. Unfavorable mutations in the complementary determining regions introduce a strain in the native protein structure which yields partial unfolding. Driven by electrostatic interactions, the protein converts into a high molecular weight, oligomeric, molten globule. The high local concentration of aggregation prone regions in the oligomer finally catalyzes the conversion into fibrils. The topology is determined by balanced electrostatic interactions in the fibril core implying a 180° rotational switch of the beta-sheets around the conserved disulfide bond.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Seeded fibrils of the germline variant of human λ-III immunoglobulin light chain FOR005 have a similar core as patient fibrils with reduced stability.J Biol Chem. 2020 Dec 25;295(52):18474-18484. doi: 10.1074/jbc.RA120.016006. Epub 2020 Oct 22. J Biol Chem. 2020. PMID: 33093170 Free PMC article.
-
Solid state NMR assignments of a human λ-III immunoglobulin light chain amyloid fibril.Biomol NMR Assign. 2021 Apr;15(1):9-16. doi: 10.1007/s12104-020-09975-2. Epub 2020 Sep 18. Biomol NMR Assign. 2021. PMID: 32946005 Free PMC article.
-
Cryo-EM reveals structural breaks in a patient-derived amyloid fibril from systemic AL amyloidosis.Nat Commun. 2021 Feb 8;12(1):875. doi: 10.1038/s41467-021-21126-2. Nat Commun. 2021. PMID: 33558536 Free PMC article.
-
Dynamic protein structures in normal function and pathologic misfolding in systemic amyloidosis.Biophys Chem. 2022 Jan;280:106699. doi: 10.1016/j.bpc.2021.106699. Epub 2021 Oct 14. Biophys Chem. 2022. PMID: 34773861 Free PMC article. Review.
-
Methods to study the structure of misfolded protein states in systemic amyloidosis.Biochem Soc Trans. 2021 Apr 30;49(2):977-985. doi: 10.1042/BST20201022. Biochem Soc Trans. 2021. PMID: 33929491 Review.
Cited by
-
Modeling immunoglobulin light chain amyloidosis in Caenorhabditis elegans.Dis Model Mech. 2025 Jul 1;18(7):dmm052230. doi: 10.1242/dmm.052230. Epub 2025 Jul 25. Dis Model Mech. 2025. PMID: 40709584 Free PMC article.
-
A conformational fingerprint for amyloidogenic light chains.Elife. 2025 Mar 3;13:RP102002. doi: 10.7554/eLife.102002. Elife. 2025. PMID: 40028903 Free PMC article.
-
Investigating Local Sequence-Structural Attributes of Amyloidogenic Light Chain Variable Domains.Proteins. 2025 Sep;93(9):1451-1464. doi: 10.1002/prot.26815. Epub 2025 Mar 4. Proteins. 2025. PMID: 40034034 Free PMC article.
-
Kinetic evidence for multiple aggregation pathways in antibody light chain variable domains.Protein Sci. 2024 Mar;33(3):e4871. doi: 10.1002/pro.4871. Protein Sci. 2024. PMID: 38100259 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
