

Engineered circular guide RNAs boost CRISPR/Cas12a- and CRISPR/Cas13d-based DNA and RNA editing
- PMID: 37353840
- PMCID: PMC10288759
- DOI: 10.1186/s13059-023-02992-z
Engineered circular guide RNAs boost CRISPR/Cas12a- and CRISPR/Cas13d-based DNA and RNA editing
Abstract
Background: The CRISPR/Cas12a and CRISPR/Cas13d systems are widely used for fundamental research and hold great potential for future clinical applications. However, the short half-life of guide RNAs (gRNAs), particularly free gRNAs without Cas nuclease binding, limits their editing efficiency and durability.
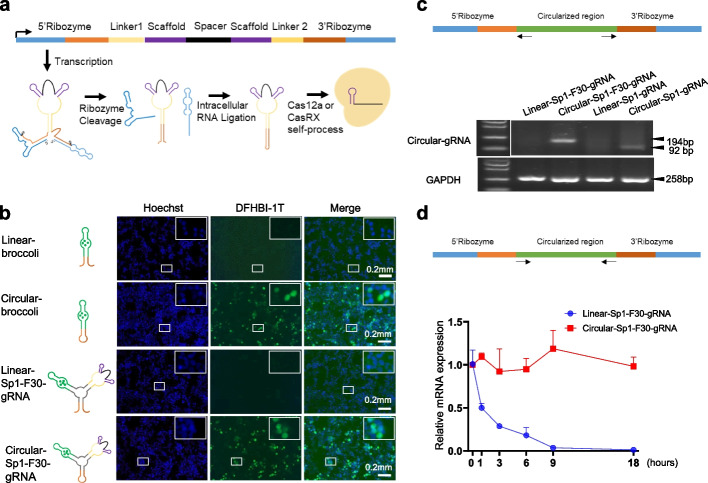
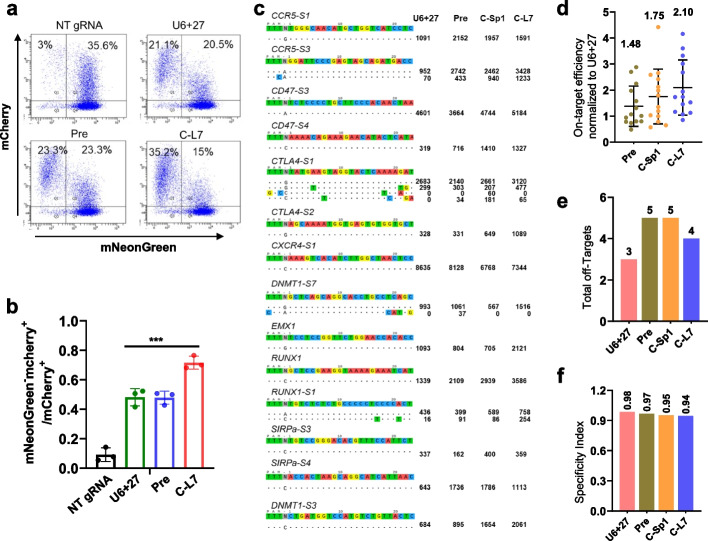
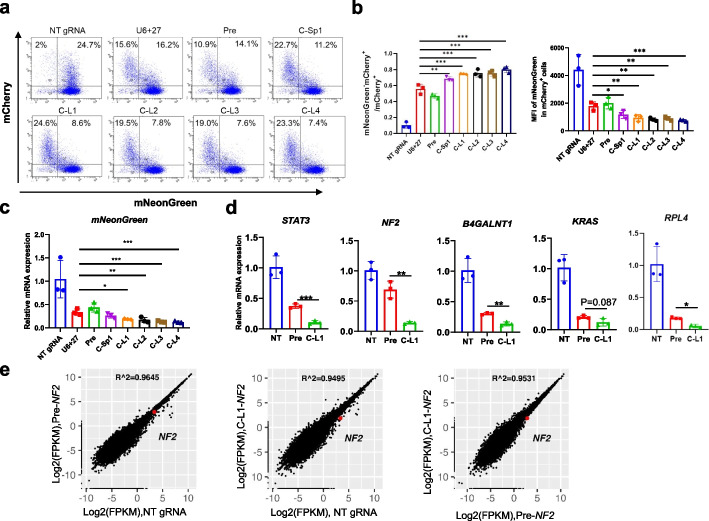
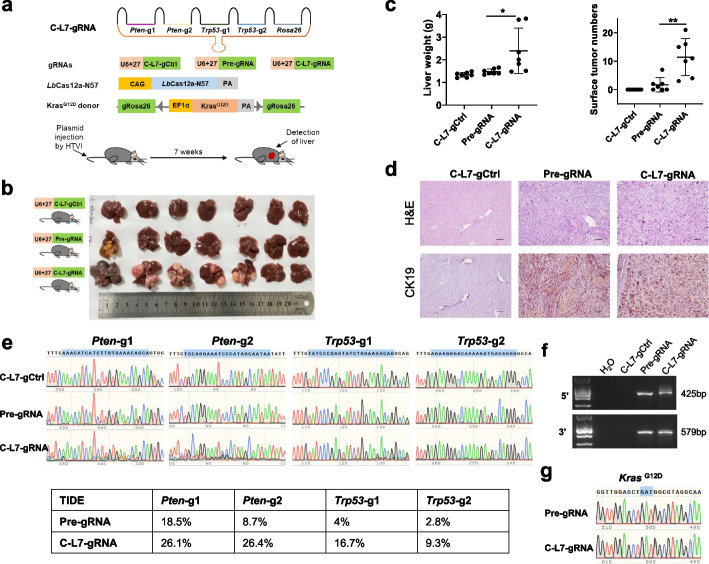
Results: Here, we engineer circular free gRNAs (cgRNAs) to increase their stability, and thus availability for Cas12a and Cas13d processing and loading, to boost editing. cgRNAs increases the efficiency of Cas12a-based transcription activators and genomic DNA cleavage by approximately 2.1- to 40.2-fold for single gene editing and 1.7- to 2.1-fold for multiplexed gene editing than their linear counterparts, without compromising specificity, across multiple sites and cell lines. Similarly, the RNA interference efficiency of Cas13d is increased by around 1.8-fold. In in vivo mouse liver, cgRNAs are more potent in activating gene expression and cleaving genomic DNA.
Conclusions: CgRNAs enable more efficient programmable DNA and RNA editing for Cas12a and Cas13d with broad applicability for fundamental research and gene therapy.
Keywords: Cas12a; Cas13d; DNA editing; Engineered circular gRNA; Gene activation; RNA editing; cgRNA.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
- 82070002/National Natural Science Foundation of China
- 82072329/National Natural Science Foundation of China
- 82102443/National Natural Science Foundation of China
- 82200072/National Natural Science Foundation of China
- 2021A1515110878/GuangDong Basic and Applied Basic Research Foundation
- 2021B1515140031/GuangDong Basic and Applied Basic Research Foundation
- 2021M701634/Fellowship of China Postdoctoral Science Foundation
- 2021M691473/Fellowship of China Postdoctoral Science Foundation
- 2022T150300/Fellowship of China Postdoctoral Science Foundation
- 2022YFA0806300/National Key R&D Program of China
- 2022YFA0806300/National Key R&D Program of China
- 2023A1515012269/GuangDong Basic and Applied Basic Research Foundation
- 2022A1515011091/GuangDong Basic and Applied Basic Research Foundation
- 2022A1515111046/GuangDong Basic and Applied Basic Research Foundation
LinkOut - more resources
Full Text Sources
Research Materials
