

A Cre Driver Line for Genetic Targeting of Kappa Opioid Receptor Expressing Cells
- PMID: 37364995
- PMCID: PMC10348446
- DOI: 10.1523/ENEURO.0043-23.2023
A Cre Driver Line for Genetic Targeting of Kappa Opioid Receptor Expressing Cells
Abstract
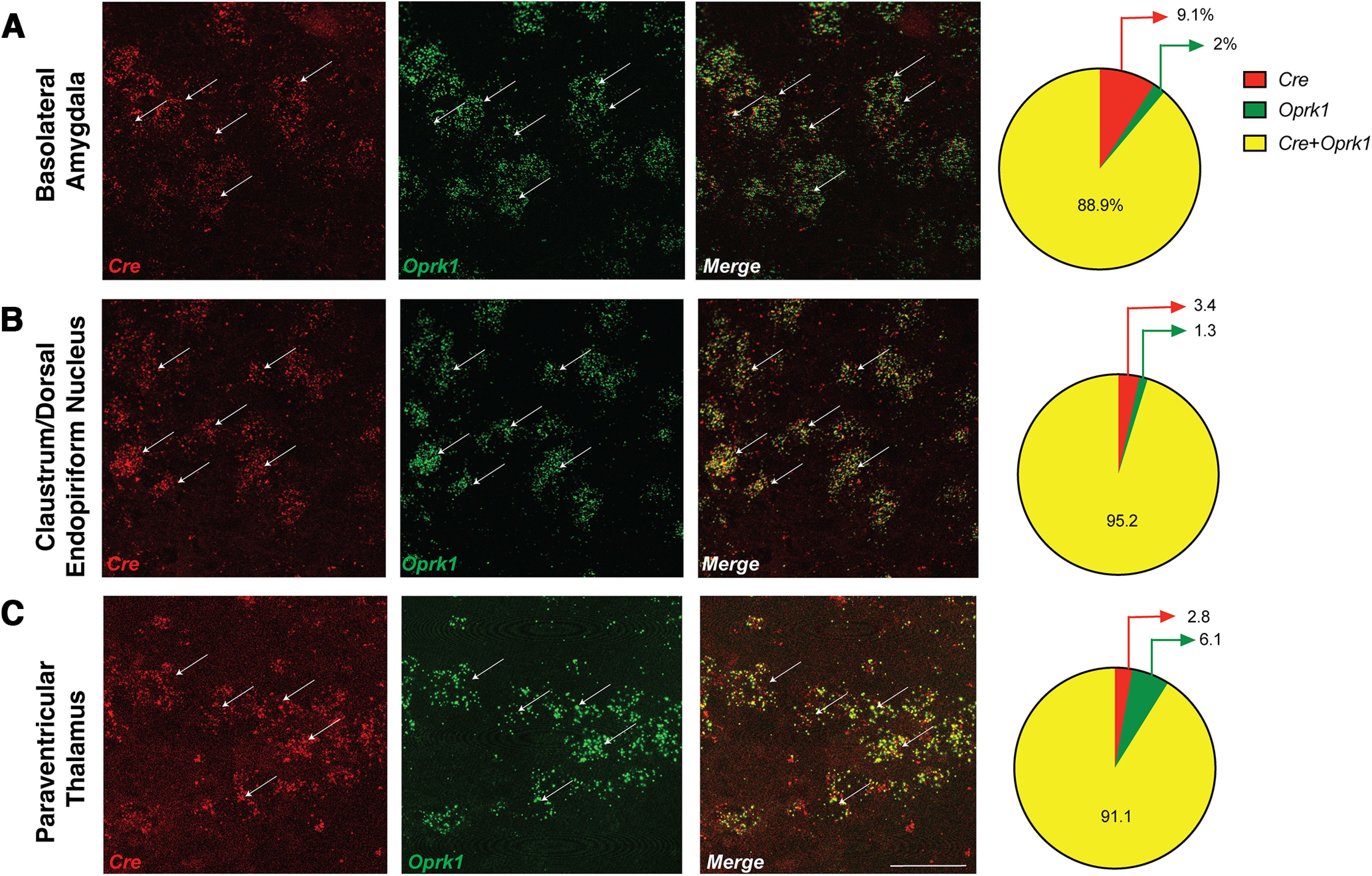
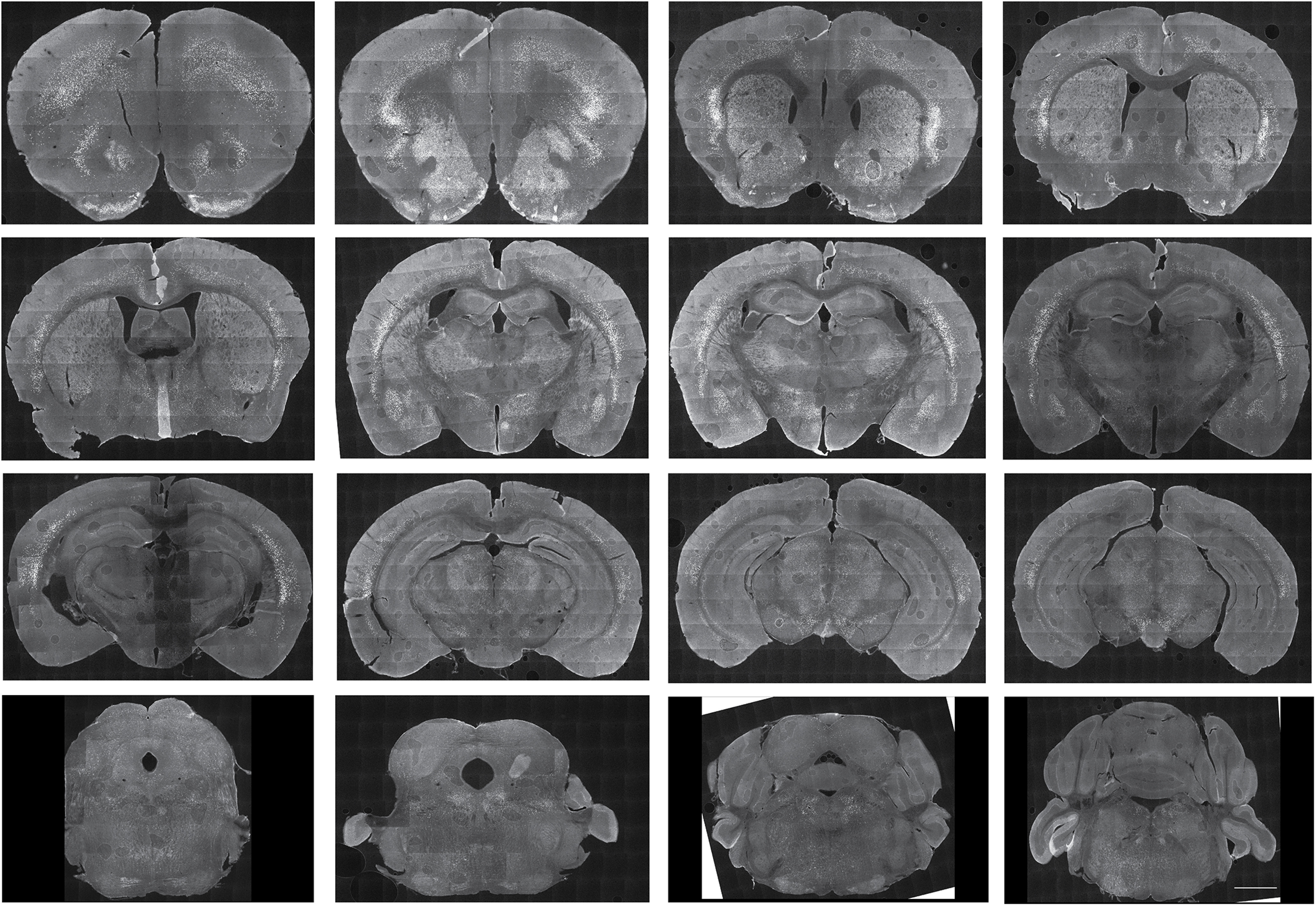
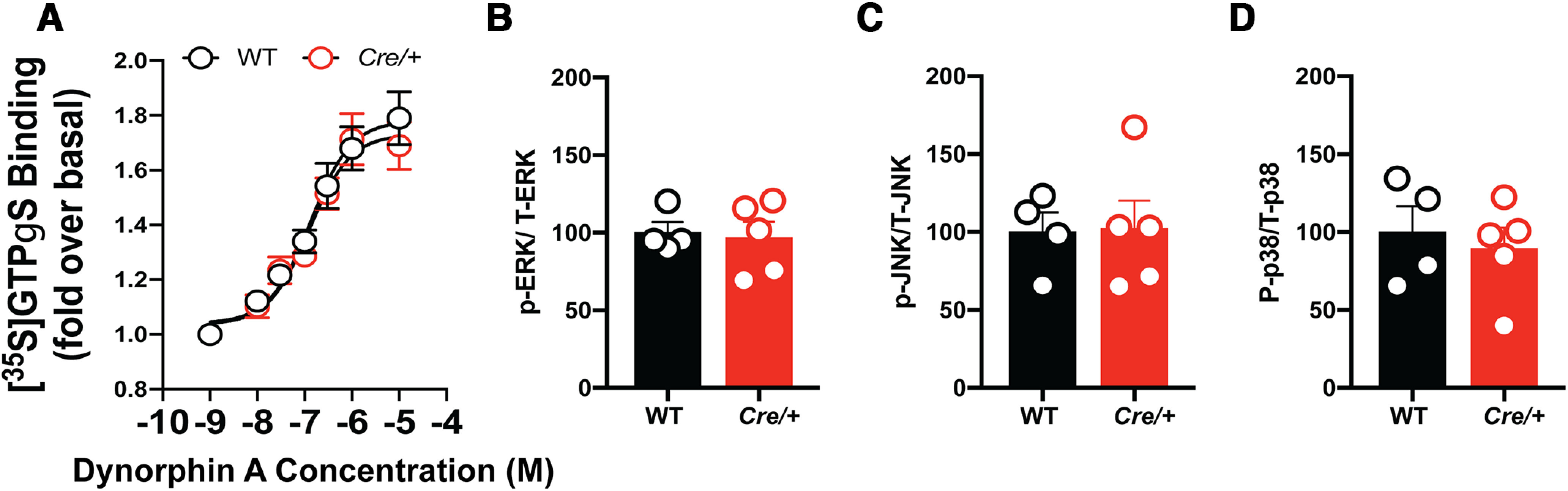
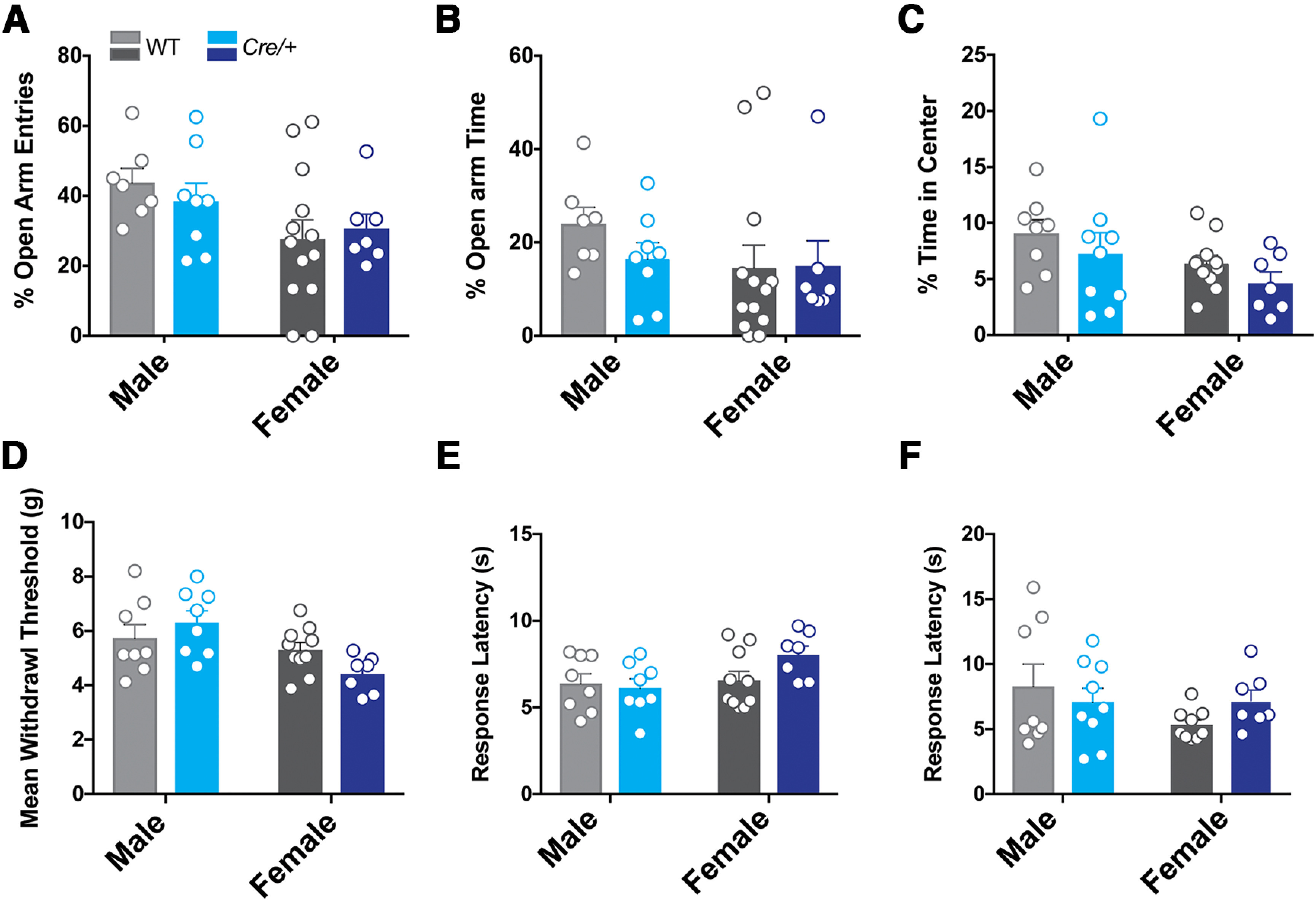
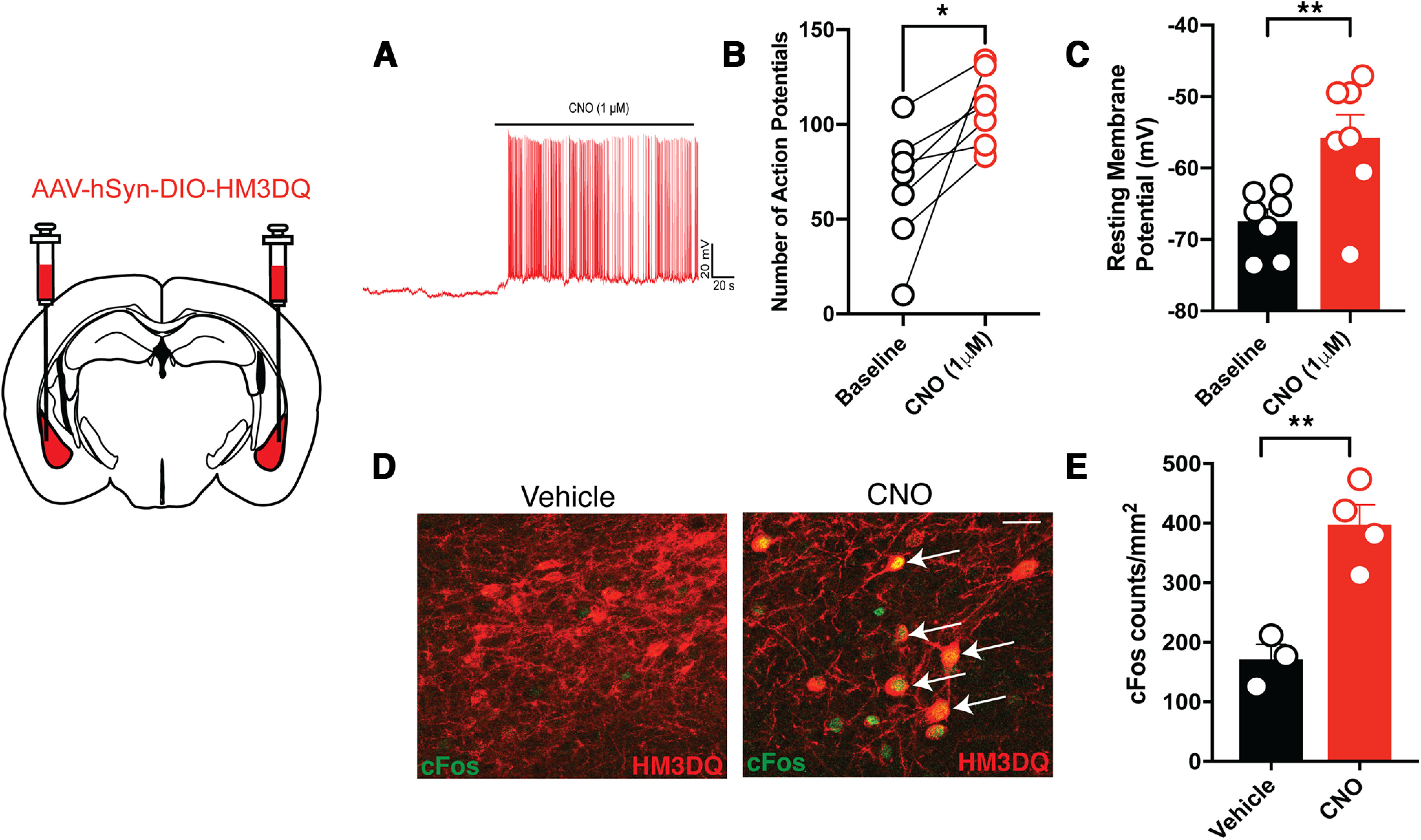
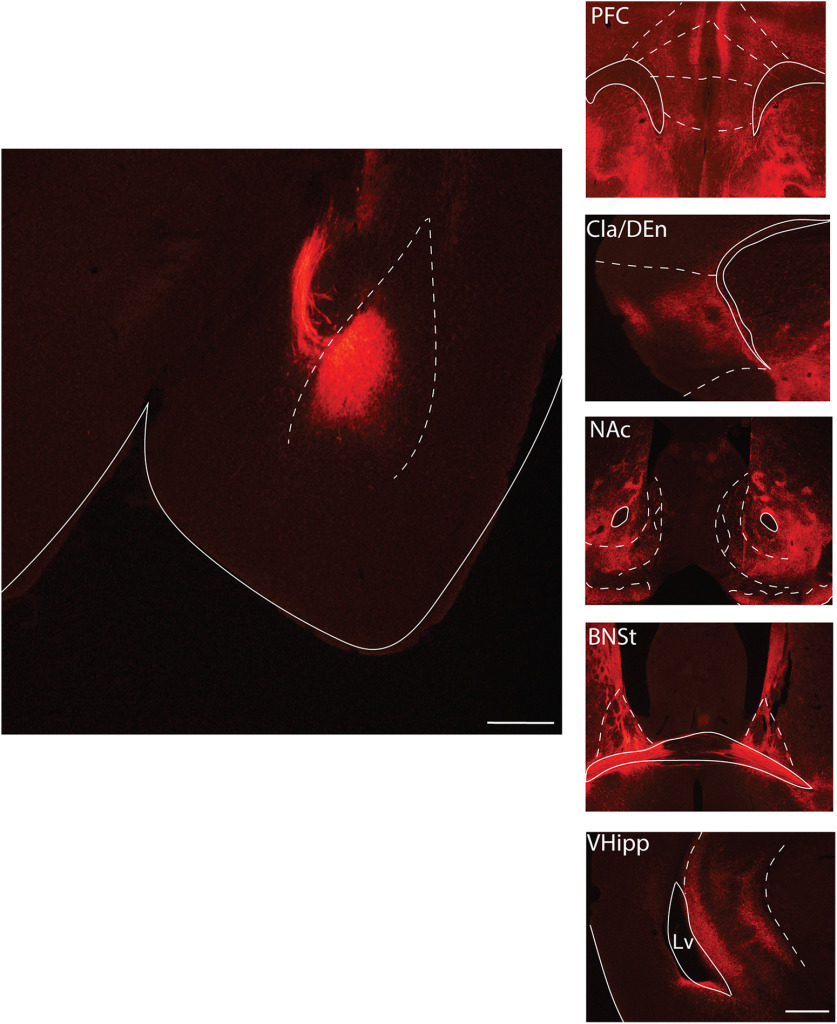
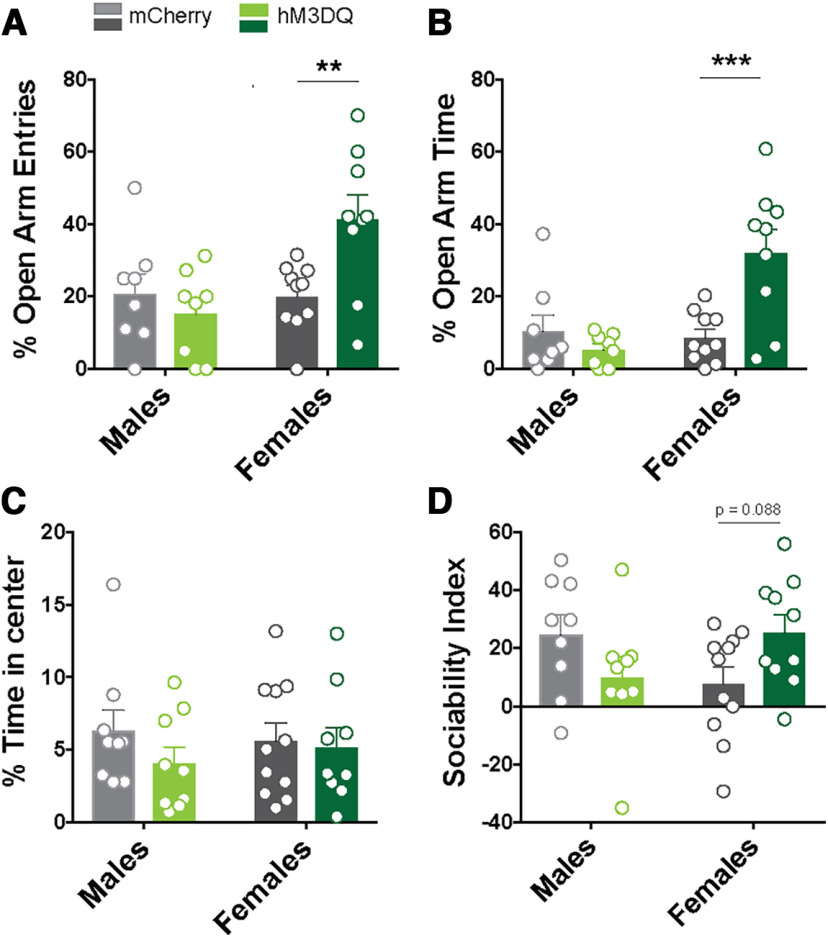
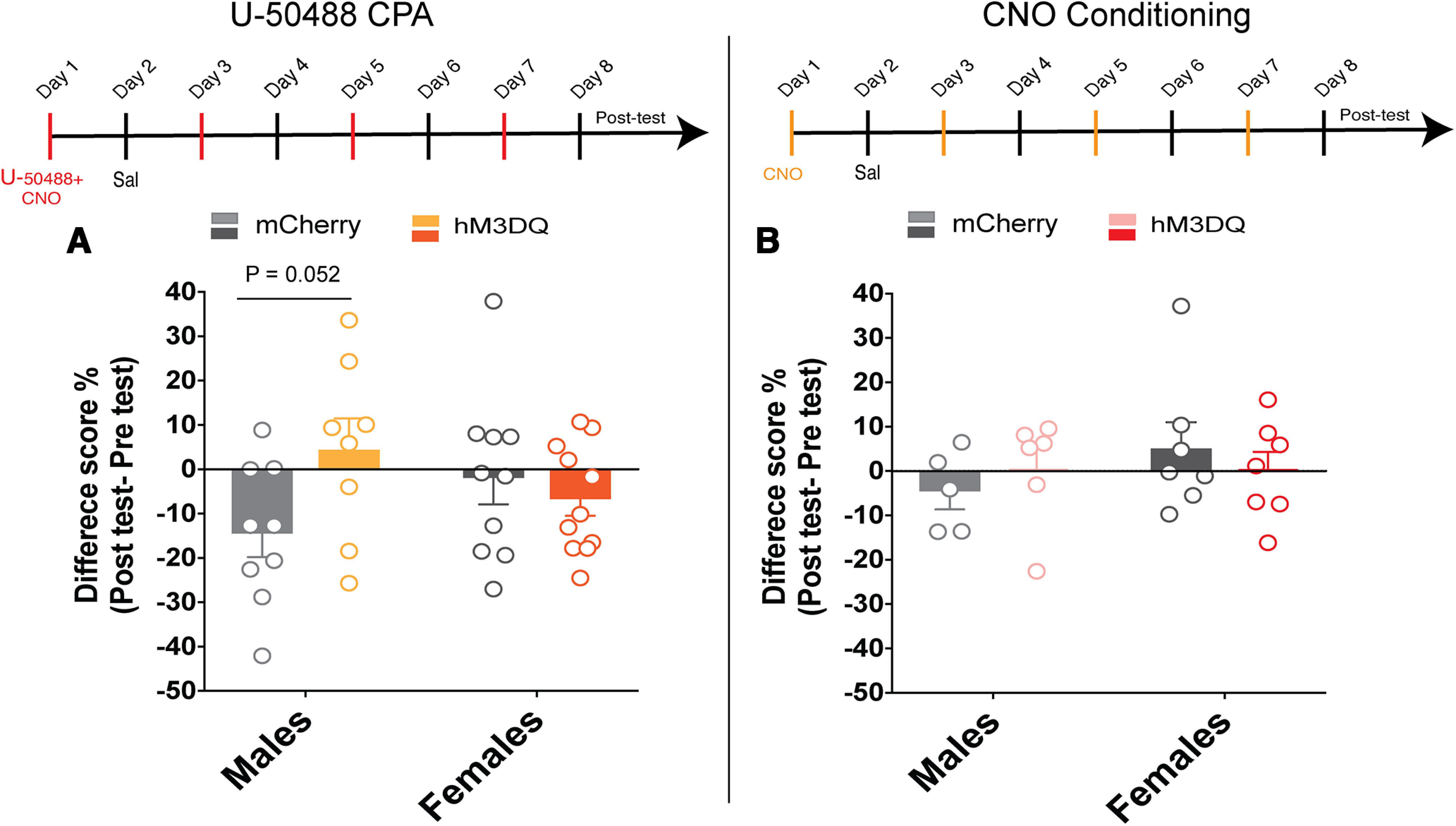
Here we describe the generation and characterization of a Cre knock-in mouse line that harbors a Cre insertion in the 3'UTR of the κ opioid receptor gene (Oprk1) locus and provides genetic access to populations of κ opioid receptor (KOR)-expressing neurons throughout the brain. Using a combination of techniques including RNA in situ hybridization and immunohistochemistry, we report that Cre is expressed with high fidelity in KOR-expressing cells throughout the brain in this mouse line. We also provide evidence that Cre insertion does not alter basal KOR function. Baseline anxiety-like behaviors and nociceptive thresholds are unaltered in Oprk1-Cre mice. Chemogenetic activation of KOR-expressing cells in the basolateral amygdala (BLAKOR cells) resulted in several sex-specific effects on anxiety-like and aversive behaviors. Activation led to decreased anxiety-like behavior on the elevated plus maze and increased sociability in female but not in male Oprk1-Cre mice. Activation of BLAKOR cells also attenuated KOR agonist-induced conditioned place aversion (CPA) in male Oprk1-Cre mice. Overall, these results suggest a potential role for BLAKOR cells in regulating anxiety-like behaviors and KOR-agonist mediated CPA. In summary, these results provide evidence for the utility of the newly generated Oprk1-Cre mice in assessing localization, anatomy, and function of KOR circuits throughout the brain.
Keywords: anxiety; conditioned place aversion; dynorphin; genetic access; knock-in mice; social interaction.
Copyright © 2023 Paliarin et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases