

Dynamic phosphorylation of CENP-N by CDK1 guides accurate chromosome segregation in mitosis
- PMID: 37365681
- PMCID: PMC10799313
- DOI: 10.1093/jmcb/mjad041
Dynamic phosphorylation of CENP-N by CDK1 guides accurate chromosome segregation in mitosis
Abstract
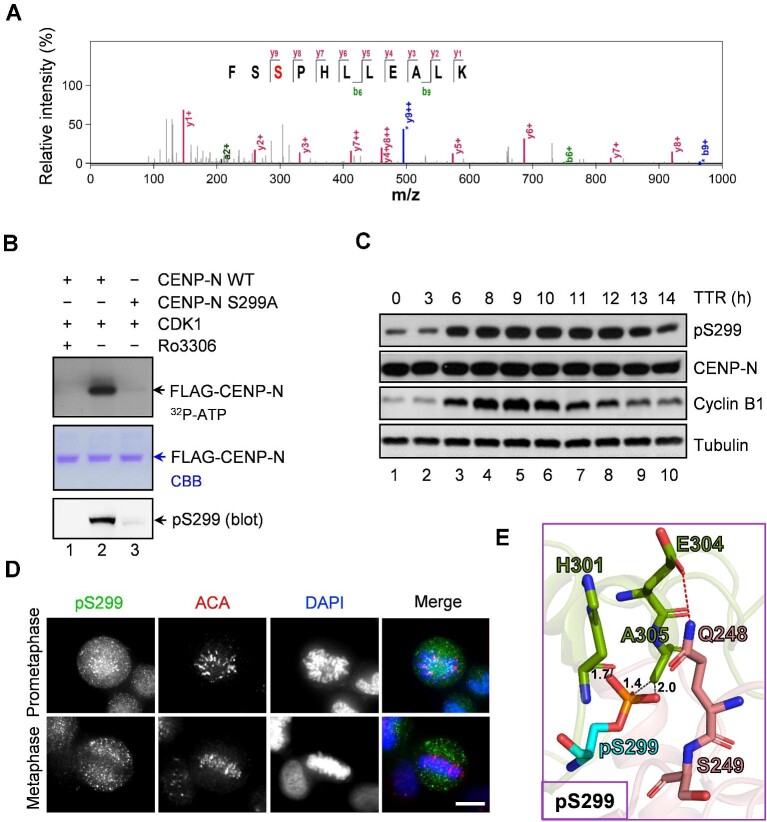
In mitosis, accurate chromosome segregation depends on the kinetochore, a supermolecular machinery that couples dynamic spindle microtubules to centromeric chromatin. However, the structure-activity relationship of the constitutive centromere-associated network (CCAN) during mitosis remains uncharacterized. Building on our recent cryo-electron microscopic analyses of human CCAN structure, we investigated how dynamic phosphorylation of human CENP-N regulates accurate chromosome segregation. Our mass spectrometric analyses revealed mitotic phosphorylation of CENP-N by CDK1, which modulates the CENP-L-CENP-N interaction for accurate chromosome segregation and CCAN organization. Perturbation of CENP-N phosphorylation is shown to prevent proper chromosome alignment and activate the spindle assembly checkpoint. These analyses provide mechanistic insight into a previously undefined link between the centromere-kinetochore network and accurate chromosome segregation.
Keywords: CDK1; CENP-N; centromere; mitosis; phosphorylation.
© The Author(s) (2023). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, CEMCS, CAS.
Figures




References
-
- Cheeseman I.M., Desai A. (2008). Molecular architecture of the kinetochore–microtubule interface. Nat. Rev. Mol. Cell Biol. 9, 33–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
