

Gamma secretase activating protein promotes end-organ dysfunction after bacterial pneumonia
- PMID: 37366533
- PMCID: PMC10396227
- DOI: 10.1152/ajplung.00018.2023
Gamma secretase activating protein promotes end-organ dysfunction after bacterial pneumonia
Abstract
Pneumonia elicits the production of cytotoxic beta amyloid (Aβ) that contributes to end-organ dysfunction, yet the mechanism(s) linking infection to activation of the amyloidogenic pathway that produces cytotoxic Aβ is unknown. Here, we tested the hypothesis that gamma-secretase activating protein (GSAP), which contributes to the amyloidogenic pathway in the brain, promotes end-organ dysfunction following bacterial pneumonia. First-in-kind Gsap knockout rats were generated. Wild-type and knockout rats possessed similar body weights, organ weights, circulating blood cell counts, arterial blood gases, and cardiac indices at baseline. Intratracheal Pseudomonas aeruginosa infection caused acute lung injury and a hyperdynamic circulatory state. Whereas infection led to arterial hypoxemia in wild-type rats, the alveolar-capillary barrier integrity was preserved in Gsap knockout rats. Infection potentiated myocardial infarction following ischemia-reperfusion injury, and this potentiation was abolished in knockout rats. In the hippocampus, GSAP contributed to both pre- and postsynaptic neurotransmission, increasing the presynaptic action potential recruitment, decreasing neurotransmitter release probability, decreasing the postsynaptic response, and preventing postsynaptic hyperexcitability, resulting in greater early long-term potentiation but reduced late long-term potentiation. Infection abolished early and late long-term potentiation in wild-type rats, whereas the late long-term potentiation was partially preserved in Gsap knockout rats. Furthermore, hippocampi from knockout rats, and both the wild-type and knockout rats following infection, exhibited a GSAP-dependent increase in neurotransmitter release probability and postsynaptic hyperexcitability. These results elucidate an unappreciated role for GSAP in innate immunity and highlight the contribution of GSAP to end-organ dysfunction during infection.NEW & NOTEWORTHY Pneumonia is a common cause of end-organ dysfunction, both during and in the aftermath of infection. In particular, pneumonia is a common cause of lung injury, increased risk of myocardial infarction, and neurocognitive dysfunction, although the mechanisms responsible for such increased risk are unknown. Here, we reveal that gamma-secretase activating protein, which contributes to the amyloidogenic pathway, is important for end-organ dysfunction following infection.
Keywords: acute lung injury; beta amyloid; long-term potentiation; myocardial infarction; pneumonia.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures






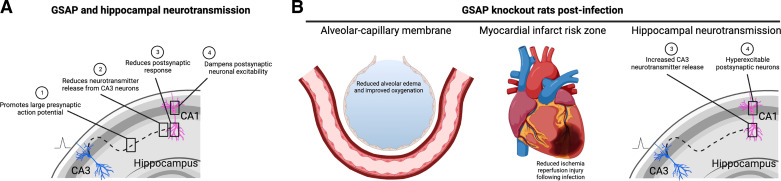
 the presynaptic depolarizing current. B: the Gsap knockout rats exhibited improved lung and heart function following infection. In lung, the alveolar-capillary membrane was protected in the knockout animals. In heart, Gsap knockout animals were not protected from ischemia-reperfusion injury under baseline conditions, but did not exhibit the postinfection potentiation of the ischemia-reperfusion injury that was seen in wild-type animals. In the hippocampus, Gsap knockout rats exhibited increased CA3 neurotransmitter release and postsynaptic hyperexcitability following infection. GSAP, gamma-secretase activating protein. [Image created with BioRender.com and published with permission.]
the presynaptic depolarizing current. B: the Gsap knockout rats exhibited improved lung and heart function following infection. In lung, the alveolar-capillary membrane was protected in the knockout animals. In heart, Gsap knockout animals were not protected from ischemia-reperfusion injury under baseline conditions, but did not exhibit the postinfection potentiation of the ischemia-reperfusion injury that was seen in wild-type animals. In the hippocampus, Gsap knockout rats exhibited increased CA3 neurotransmitter release and postsynaptic hyperexcitability following infection. GSAP, gamma-secretase activating protein. [Image created with BioRender.com and published with permission.]References
-
- Chang JC, Xu P, Wong E, Flajolet M, Li YM, Greengard P. GSAP regulates amyloid β production through modulation of amyloid precursor protein trafficking. bioRxiv, 2020. doi:10.1101/2020.11.12.379313. - DOI
-
- Xu P, Chang JC, Zhou X, Wang W, Bamkole M, Wong E, Bettayeb K, Jiang LL, Huang T, Luo W, Xu H, Nairn AC, Flajolet M, Ip NY, Li YM, Greengard P. GSAP regulates lipid homeostasis and mitochondrial function associated with Alzheimer's disease. J Exp Med 218: e20202446, 2021. doi:10.1084/jem.20202446. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- R01 HL167997/HL/NHLBI NIH HHS/United States
- R01 HL160988/HL/NHLBI NIH HHS/United States
- S10 OD025089/OD/NIH HHS/United States
- P01 HL066299/HL/NHLBI NIH HHS/United States
- R01 HL118334/HL/NHLBI NIH HHS/United States
- R01 HL140182/HL/NHLBI NIH HHS/United States
- F31 HL147512/HL/NHLBI NIH HHS/United States
- R01 HL148069/HL/NHLBI NIH HHS/United States
- P01 HL116264/HL/NHLBI NIH HHS/United States
- K99 AG058780/AG/NIA NIH HHS/United States
- R21 OD026560/OD/NIH HHS/United States
- T32 HL007778/HL/NHLBI NIH HHS/United States
- R00 AG058780/AG/NIA NIH HHS/United States
- R01 OD010944/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical