

Evo Devo of the Vertebrates Integument
- PMID: 37367479
- PMCID: PMC10299021
- DOI: 10.3390/jdb11020025
Evo Devo of the Vertebrates Integument
Abstract
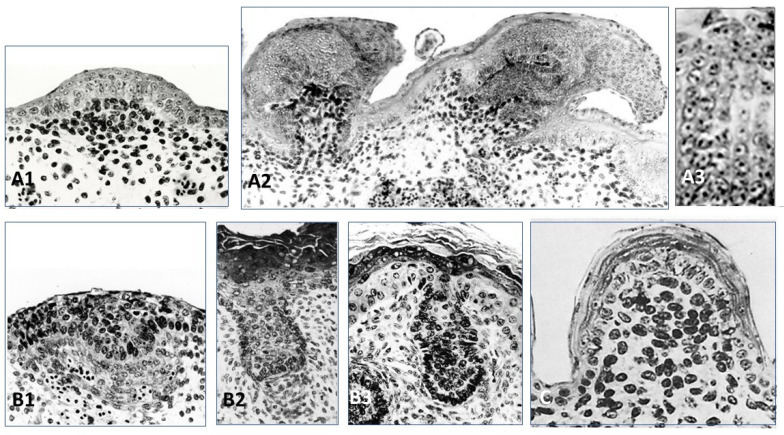
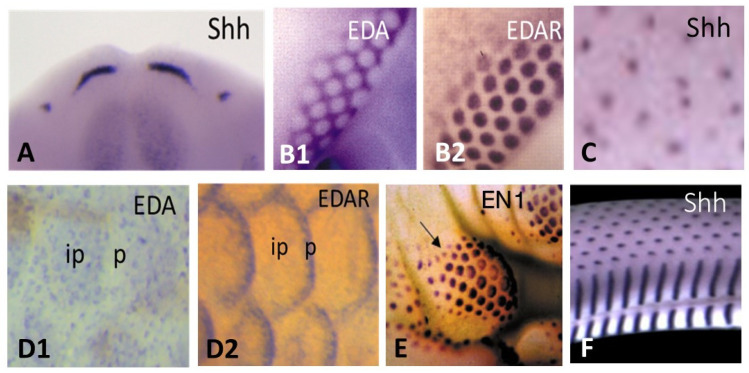
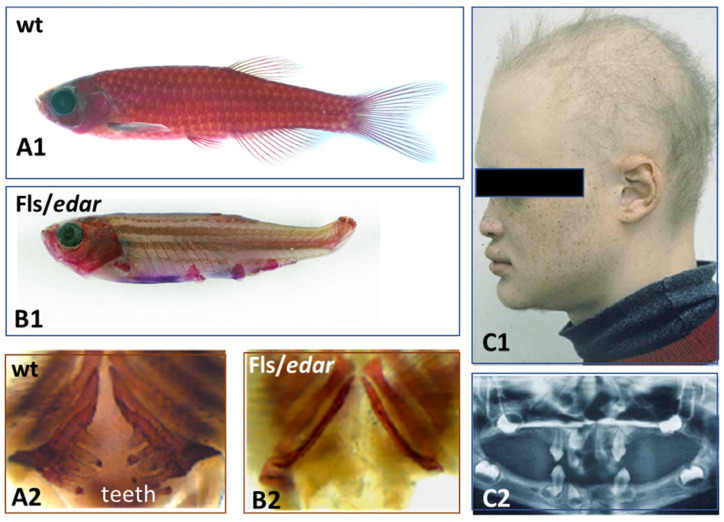
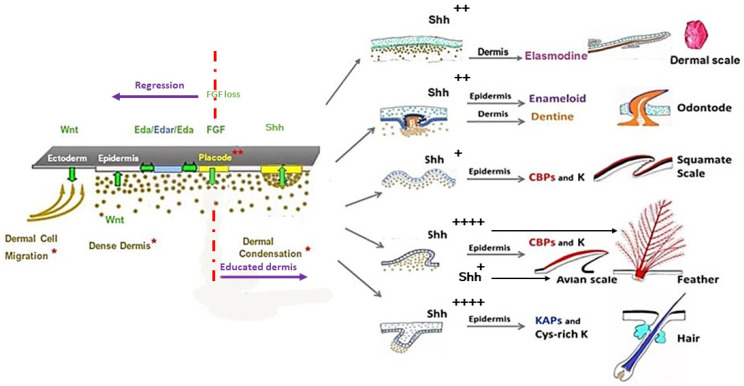
All living jawed vertebrates possess teeth or did so ancestrally. Integumental surface also includes the cornea. Conversely, no other anatomical feature differentiates the clades so readily as skin appendages do, multicellular glands in amphibians, hair follicle/gland complexes in mammals, feathers in birds, and the different types of scales. Tooth-like scales are characteristic of chondrichthyans, while mineralized dermal scales are characteristic of bony fishes. Corneous epidermal scales might have appeared twice, in squamates, and on feet in avian lineages, but posteriorly to feathers. In contrast to the other skin appendages, the origin of multicellular glands of amphibians has never been addressed. In the seventies, pioneering dermal-epidermal recombination between chick, mouse and lizard embryos showed that: (1) the clade type of the appendage is determined by the epidermis; (2) their morphogenesis requires two groups of dermal messages, first for primordia formation, second for appendage final architecture; (3) the early messages were conserved during amniotes evolution. Molecular biology studies that have identified the involved pathways, extending those data to teeth and dermal scales, suggest that the different vertebrate skin appendages evolved in parallel from a shared placode/dermal cells unit, present in a common toothed ancestor, c.a. 420 mya.
Keywords: cornea; development; evolution; feather; hair; placode; reticula; scale; tooth.
Conflict of interest statement
The author declares no conflict of interest.
Figures



Similar articles
-
Getting to the root of scales, feather and hair: As deep as odontodes?Exp Dermatol. 2019 Apr;28(4):503-508. doi: 10.1111/exd.13391. Epub 2017 Aug 25. Exp Dermatol. 2019. PMID: 28603898
-
Formation of cutaneous appendages in dermo-epidermal recombinations between reptiles, birds and mammals.Wilehm Roux Arch Dev Biol. 1975 Dec;177(4):323-340. doi: 10.1007/BF00848183. Wilehm Roux Arch Dev Biol. 1975. PMID: 28305000
-
Developing an ancient epithelial appendage: FGF signalling regulates early tail denticle formation in sharks.Evodevo. 2017 May 2;8:8. doi: 10.1186/s13227-017-0071-0. eCollection 2017. Evodevo. 2017. PMID: 28469835 Free PMC article.
-
Avian skin development and the evolutionary origin of feathers.J Exp Zool B Mol Dev Evol. 2003 Aug 15;298(1):57-72. doi: 10.1002/jez.b.26. J Exp Zool B Mol Dev Evol. 2003. PMID: 12949769 Review.
-
A new scenario for the evolutionary origin of hair, feather, and avian scales.J Anat. 2009 Apr;214(4):587-606. doi: 10.1111/j.1469-7580.2008.01041.x. J Anat. 2009. PMID: 19422430 Free PMC article. Review.
Cited by
-
A chromosome-level genome assembly of the Asian house martin implies potential genes associated with the feathered-foot trait.G3 (Bethesda). 2024 Jun 5;14(6):jkae077. doi: 10.1093/g3journal/jkae077. G3 (Bethesda). 2024. PMID: 38607414 Free PMC article.
-
Interactions between avian viruses and skin in farm birds.Vet Res. 2024 Apr 26;55(1):54. doi: 10.1186/s13567-024-01310-0. Vet Res. 2024. PMID: 38671518 Free PMC article. Review.
-
Resilience of the replacing dentition in adult reptiles.Dev Biol. 2024 Dec;516:71-81. doi: 10.1016/j.ydbio.2024.07.013. Epub 2024 Jul 24. Dev Biol. 2024. PMID: 39059678 Free PMC article. Review.
-
The origin and early evolution of feathers: implications, uncertainties and future prospects.Biol Lett. 2025 Feb;21(2):20240517. doi: 10.1098/rsbl.2024.0517. Epub 2025 Feb 19. Biol Lett. 2025. PMID: 39969251 Free PMC article. Review.
-
Development-Associated Genes of the Epidermal Differentiation Complex (EDC).J Dev Biol. 2024 Jan 15;12(1):4. doi: 10.3390/jdb12010004. J Dev Biol. 2024. PMID: 38248869 Free PMC article. Review.
References
-
- Chammas S.M., Carneiro S.M., Ferro R.S., Antoniazzi M.M., Jared C. Development of integument and cutaneous glands in larval, juvenile and adult toads (Rhinella granulosa): A morphological and morphometric study. Acta Zool. 2015;96:460–477. doi: 10.1111/azo.12091. - DOI
-
- Lucas A.M., Stettenheim P.R. Avian Anatomy Integument. US Department of Agriculture; Washington, DC, USA: 1972.
Publication types
LinkOut - more resources
Full Text Sources
