

Complement Activation-Independent Attenuation of SARS-CoV-2 Infection by C1q and C4b-Binding Protein
- PMID: 37376569
- PMCID: PMC10305604
- DOI: 10.3390/v15061269
Complement Activation-Independent Attenuation of SARS-CoV-2 Infection by C1q and C4b-Binding Protein
Abstract
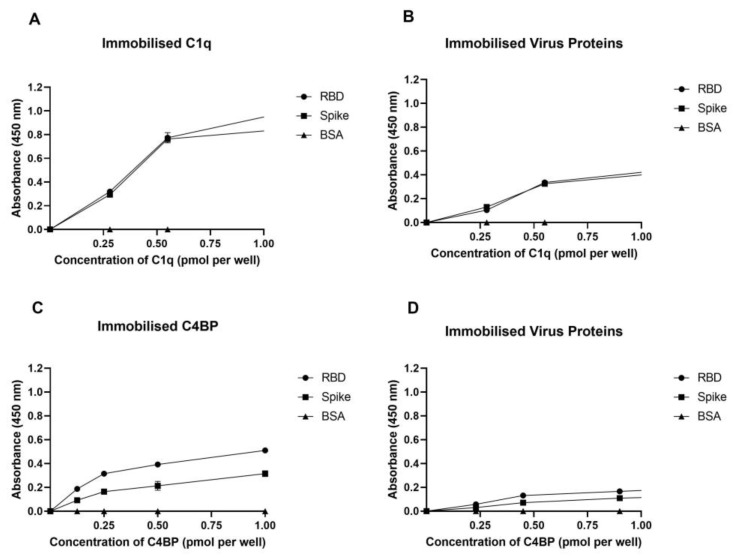
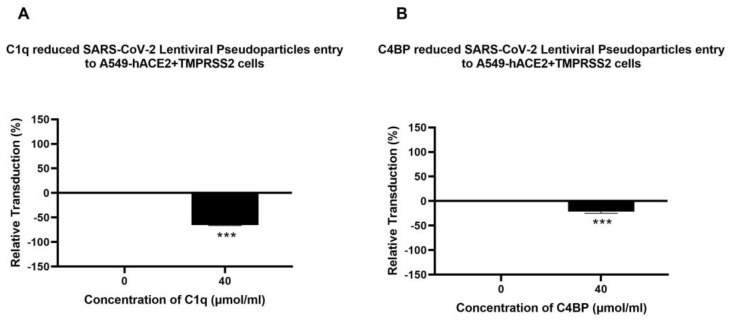
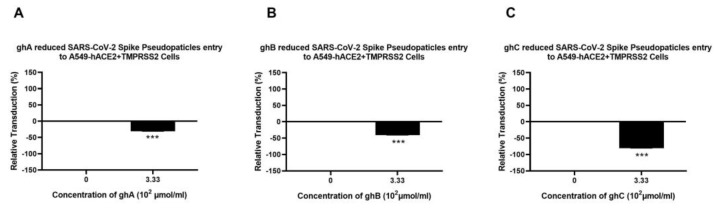
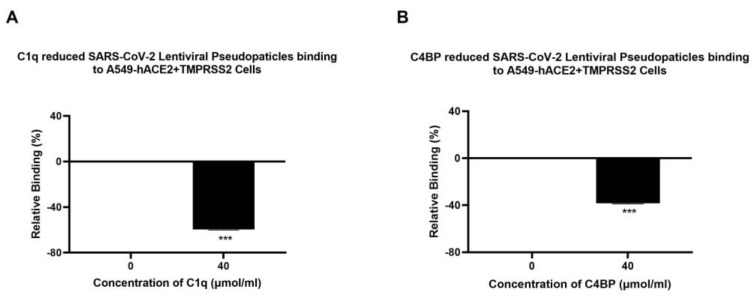
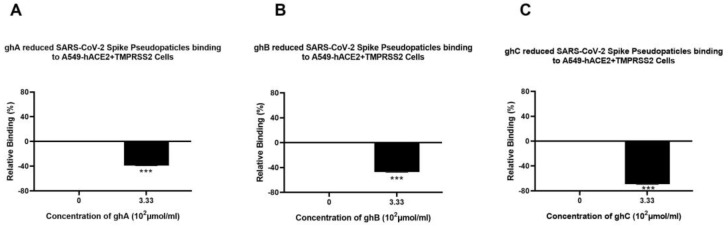
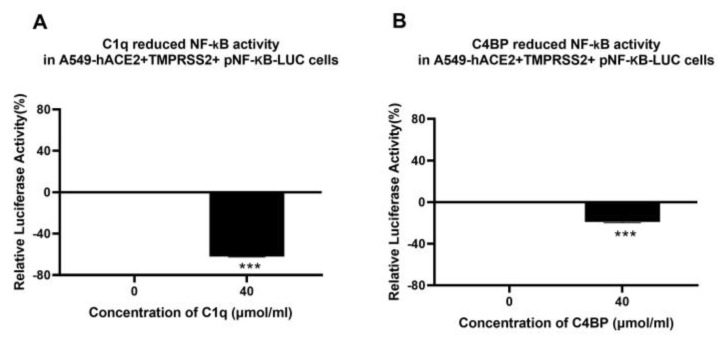
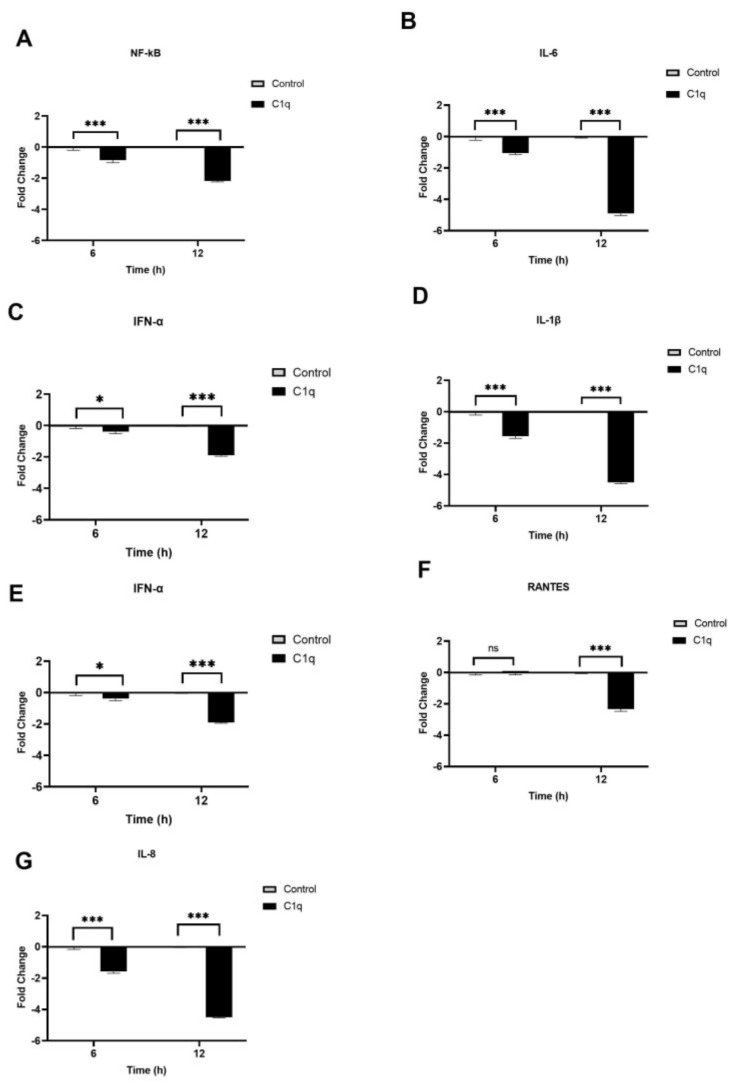
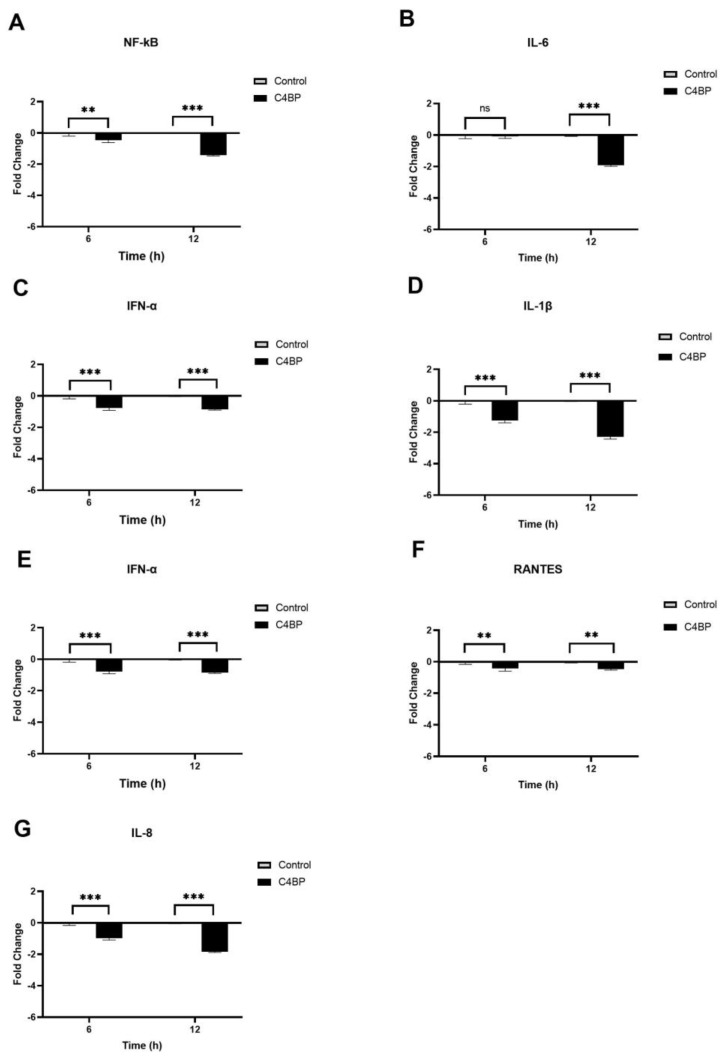
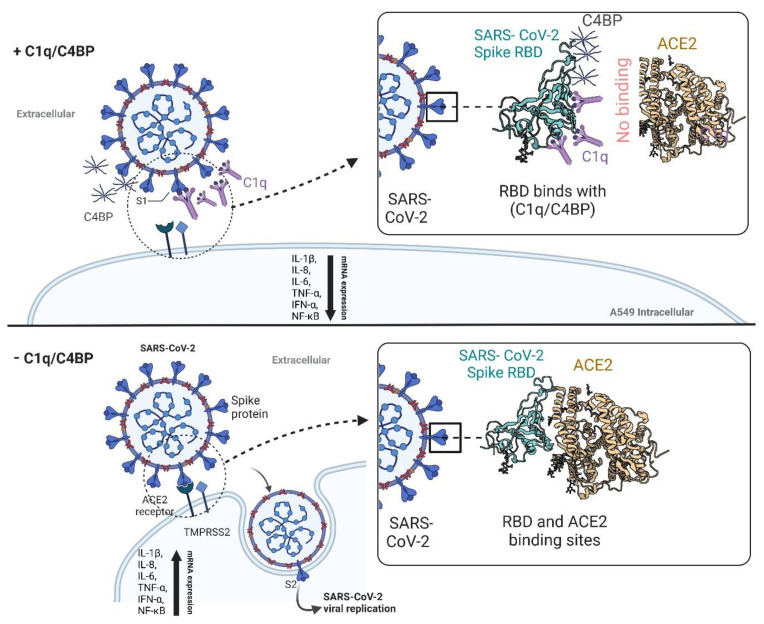
The complement system is a key component of the innate immune response to viruses and proinflammatory events. Exaggerated complement activation has been attributed to the induction of a cytokine storm in severe SARS-CoV-2 infection. However, there is also an argument for the protective role of complement proteins, given their local synthesis or activation at the site of viral infection. This study investigated the complement activation-independent role of C1q and C4b-binding protein (C4BP) against SARS-CoV-2 infection. The interactions of C1q, its recombinant globular heads, and C4BP with the SARS-CoV-2 spike and receptor binding domain (RBD) were examined using direct ELISA. In addition, RT-qPCR was used to evaluate the modulatory effect of these complement proteins on the SARS-CoV-2-mediated immune response. Cell binding and luciferase-based viral entry assays were utilised to assess the effects of C1q, its recombinant globular heads, and C4BP on SARS-CoV-2 cell entry. C1q and C4BP bound directly to SARS-CoV-2 pseudotype particles via the RBD domain of the spike protein. C1q via its globular heads and C4BP were found to reduce binding as well as viral transduction of SARS-CoV-2 spike protein expressing lentiviral pseudotypes into transfected A549 cells expressing human ACE2 and TMPRSS2. Furthermore, the treatment of the SARS-CoV-2 spike, envelope, nucleoprotein, and membrane protein expressing alphaviral pseudotypes with C1q, its recombinant globular heads, or C4BP triggered a reduction in mRNA levels of proinflammatory cytokines and chemokines such as IL-1β, IL-8, IL-6, TNF-α, IFN-α, and RANTES (as well as NF-κB) in A549 cells expressing human ACE2 and TMPRSS2. In addition, C1q and C4BP treatment also reduced SARS-CoV-2 pseudotype infection-mediated NF-κB activation in A549 cells expressing human ACE2 and TMPRSS2. C1q and C4BP are synthesised primarily by hepatocytes; however, they are also produced by macrophages, and alveolar type II cells, respectively, locally at the pulmonary site. These findings support the notion that the locally produced C1q and C4BP can be protective against SARS-CoV-2 infection in a complement activation-independent manner, offering immune resistance by inhibiting virus binding to target host cells and attenuating the infection-associated inflammatory response.
Keywords: C1q; C4BP; COVID-19; SARS-CoV-2; classical pathway; complement; innate immunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cascella M., Rajnik M., Aleem A., Dulebohn S.C., Di Napoli R. StatPearls [Internet] StatPearls; Treasure Island, FL, USA: 2022. Features, evaluation, and treatment of coronavirus (COVID-19) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
