

A standardized quantitative analysis strategy for stable isotope probing metagenomics
- PMID: 37377419
- PMCID: PMC10469821
- DOI: 10.1128/msystems.01280-22
A standardized quantitative analysis strategy for stable isotope probing metagenomics
Abstract
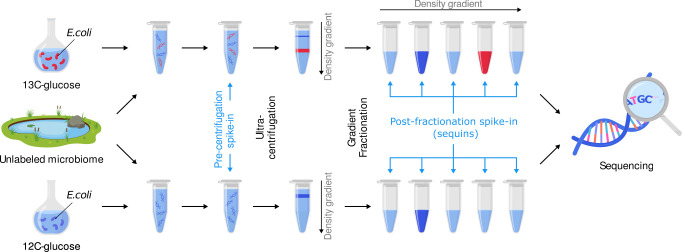
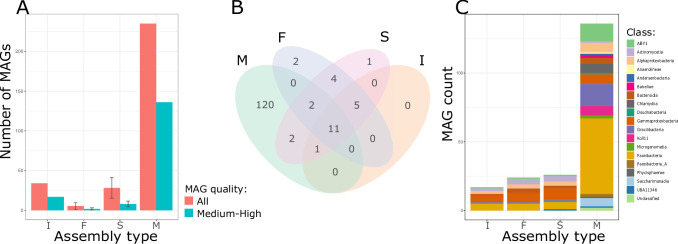
Stable isotope probing (SIP) facilitates culture-independent identification of active microbial populations within complex ecosystems through isotopic enrichment of nucleic acids. Many DNA-SIP studies rely on 16S rRNA gene sequences to identify active taxa, but connecting these sequences to specific bacterial genomes is often challenging. Here, we describe a standardized laboratory and analysis framework to quantify isotopic enrichment on a per-genome basis using shotgun metagenomics instead of 16S rRNA gene sequencing. To develop this framework, we explored various sample processing and analysis approaches using a designed microbiome where the identity of labeled genomes and their level of isotopic enrichment were experimentally controlled. With this ground truth dataset, we empirically assessed the accuracy of different analytical models for identifying active taxa and examined how sequencing depth impacts the detection of isotopically labeled genomes. We also demonstrate that using synthetic DNA internal standards to measure absolute genome abundances in SIP density fractions improves estimates of isotopic enrichment. In addition, our study illustrates the utility of internal standards to reveal anomalies in sample handling that could negatively impact SIP metagenomic analyses if left undetected. Finally, we present SIPmg, an R package to facilitate the estimation of absolute abundances and perform statistical analyses for identifying labeled genomes within SIP metagenomic data. This experimentally validated analysis framework strengthens the foundation of DNA-SIP metagenomics as a tool for accurately measuring the in situ activity of environmental microbial populations and assessing their genomic potential. IMPORTANCE Answering the questions, "who is eating what?" and "who is active?" within complex microbial communities is paramount for our ability to model, predict, and modulate microbiomes for improved human and planetary health. These questions can be pursued using stable isotope probing to track the incorporation of labeled compounds into cellular DNA during microbial growth. However, with traditional stable isotope methods, it is challenging to establish links between an active microorganism's taxonomic identity and genome composition while providing quantitative estimates of the microorganism's isotope incorporation rate. Here, we report an experimental and analytical workflow that lays the foundation for improved detection of metabolically active microorganisms and better quantitative estimates of genome-resolved isotope incorporation, which can be used to further refine ecosystem-scale models for carbon and nutrient fluxes within microbiomes.
Keywords: DNA-SIP; co-assembly; internal standards; metagenomics; spike-ins; stable isotope probing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
