

The Life Cycle of Aurelia aurita Depends on the Presence of a Microbiome in Polyps Prior to Onset of Strobilation
- PMID: 37378516
- PMCID: PMC10433978
- DOI: 10.1128/spectrum.00262-23
The Life Cycle of Aurelia aurita Depends on the Presence of a Microbiome in Polyps Prior to Onset of Strobilation
Abstract
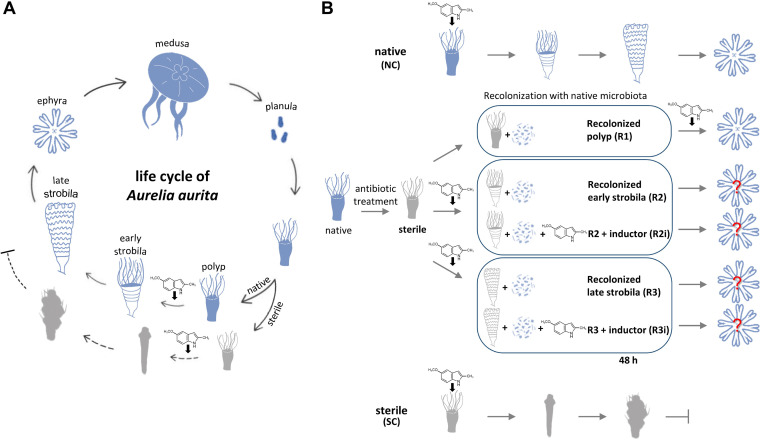
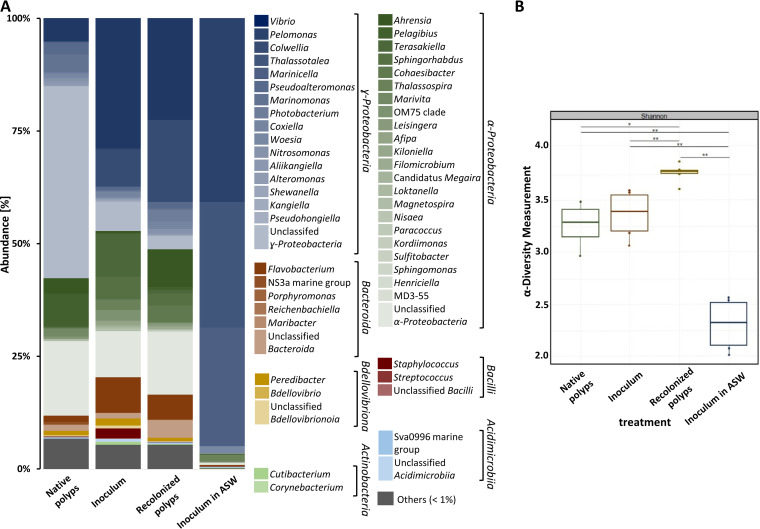
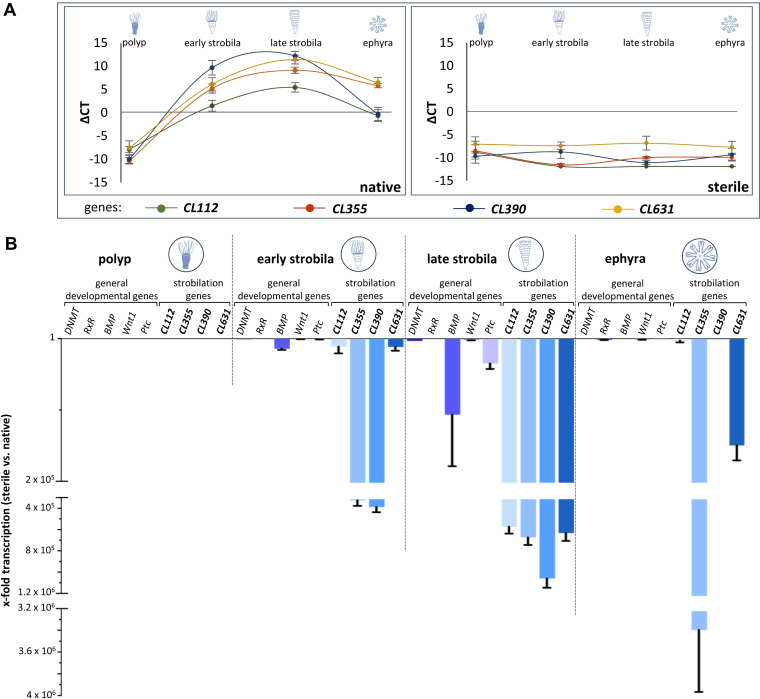
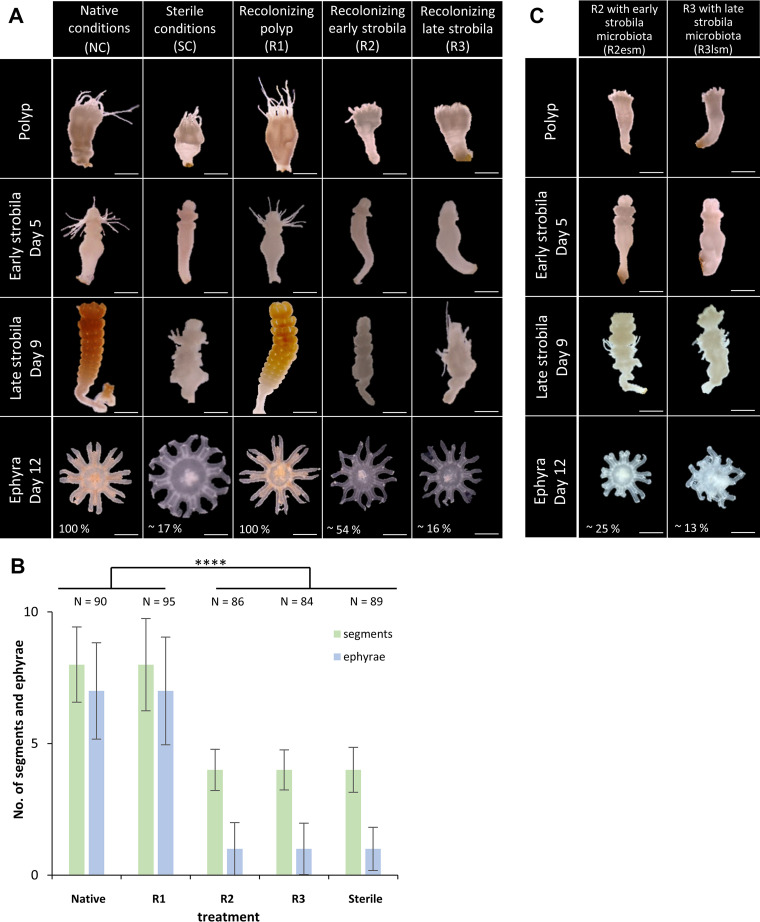
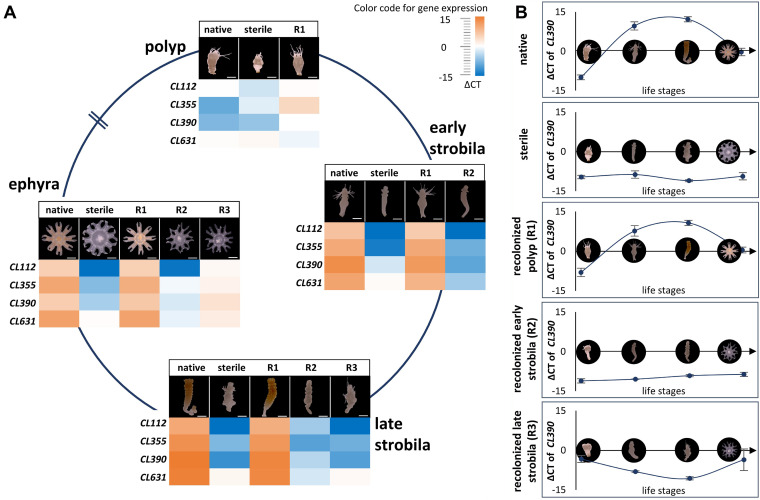
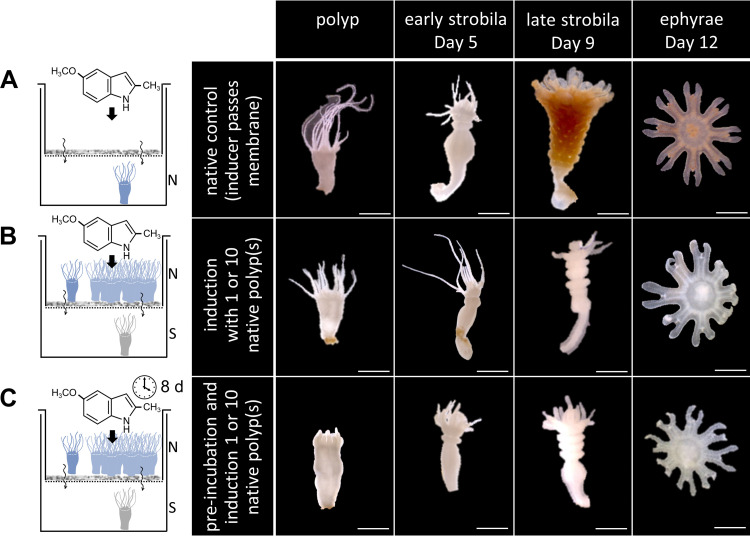
Aurelia aurita's intricate life cycle alternates between benthic polyp and pelagic medusa stages. The strobilation process, a critical asexual reproduction mechanism in this jellyfish, is severely compromised in the absence of the natural polyp microbiome, with limited production and release of ephyrae. Yet, the recolonization of sterile polyps with a native polyp microbiome can correct this defect. Here, we investigated the precise timing necessary for recolonization as well as the host-associated molecular processes involved. We deciphered that a natural microbiota had to be present in polyps prior to the onset of strobilation to ensure normal asexual reproduction and a successful polyp-to-medusa transition. Providing the native microbiota to sterile polyps after the onset of strobilation failed to restore the normal strobilation process. The absence of a microbiome was associated with decreased transcription of developmental and strobilation genes as monitored by reverse transcription-quantitative PCR. Transcription of these genes was exclusively observed for native polyps and sterile polyps that were recolonized before the initiation of strobilation. We further propose that direct cell contact between the host and its associated bacteria is required for the normal production of offspring. Overall, our findings indicate that the presence of a native microbiome at the polyp stage prior to the onset of strobilation is essential to ensure a normal polyp-to-medusa transition. IMPORTANCE All multicellular organisms are associated with microorganisms that play fundamental roles in the health and fitness of the host. Notably, the native microbiome of the Cnidarian Aurelia aurita is crucial for the asexual reproduction by strobilation. Sterile polyps display malformed strobilae and a halt of ephyrae release, which is restored by recolonizing sterile polyps with a native microbiota. Despite that, little is known about the microbial impact on the strobilation process's timing and molecular consequences. The present study shows that A. aurita's life cycle depends on the presence of the native microbiome at the polyp stage prior to the onset of strobilation to ensure the polyp-to-medusa transition. Moreover, sterile individuals correlate with reduced transcription levels of developmental and strobilation genes, evidencing the microbiome's impact on strobilation on the molecular level. Transcription of strobilation genes was exclusively detected in native polyps and those recolonized before initiating strobilation, suggesting microbiota-dependent gene regulation.
Keywords: Aurelia aurita; asexual reproduction; life cycle; native microbiota; recolonization; reproduction; strobilation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Unabia C, Hadfield M. 1999. Role of bacteria in larval settlement and metamorphosis of the polychaete Hydroides elegans. Marine Biology 133:55–64. doi: 10.1007/s002270050442. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
