

The diverse structural modes of tRNA binding and recognition
- PMID: 37380076
- PMCID: PMC10424219
- DOI: 10.1016/j.jbc.2023.104966
The diverse structural modes of tRNA binding and recognition
Abstract
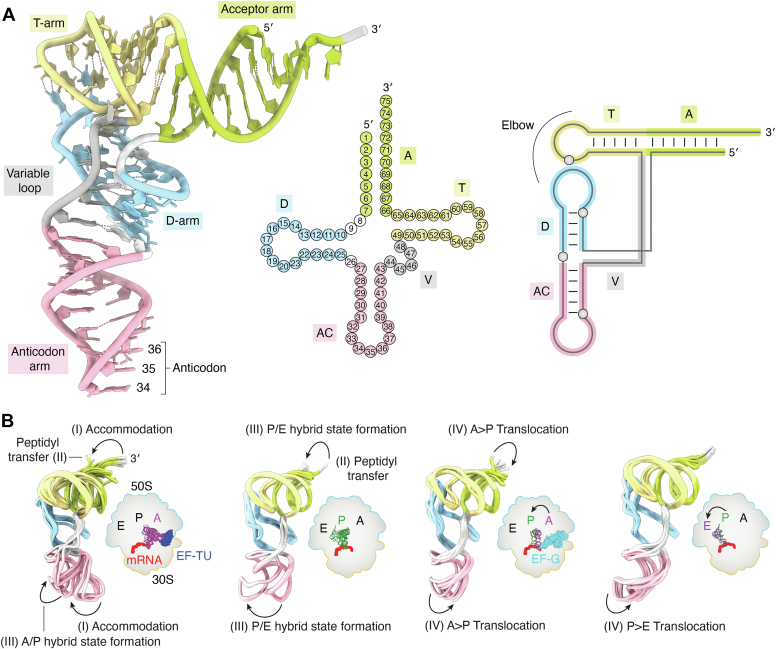
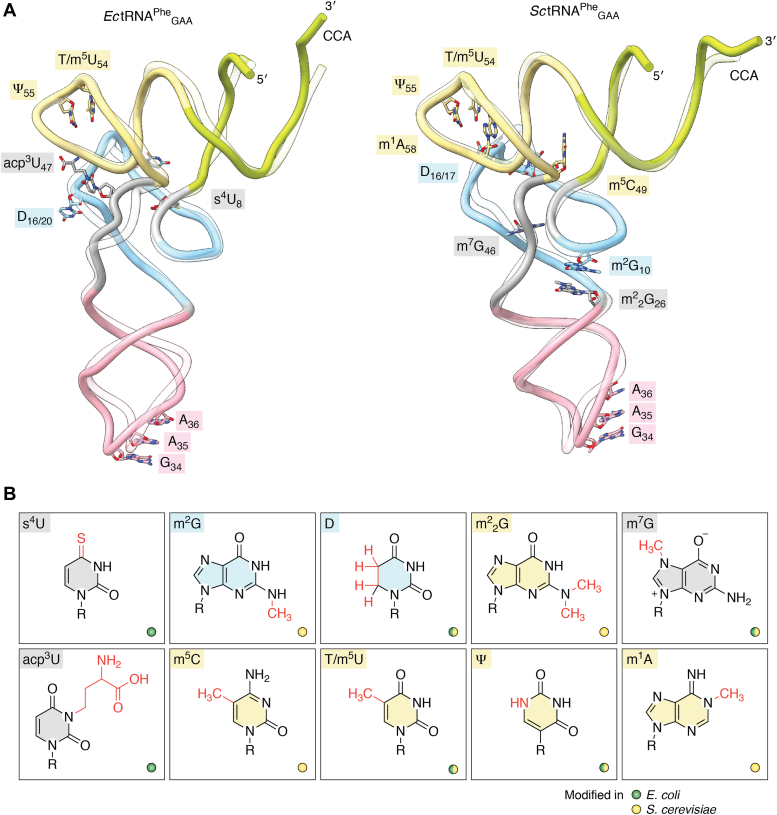
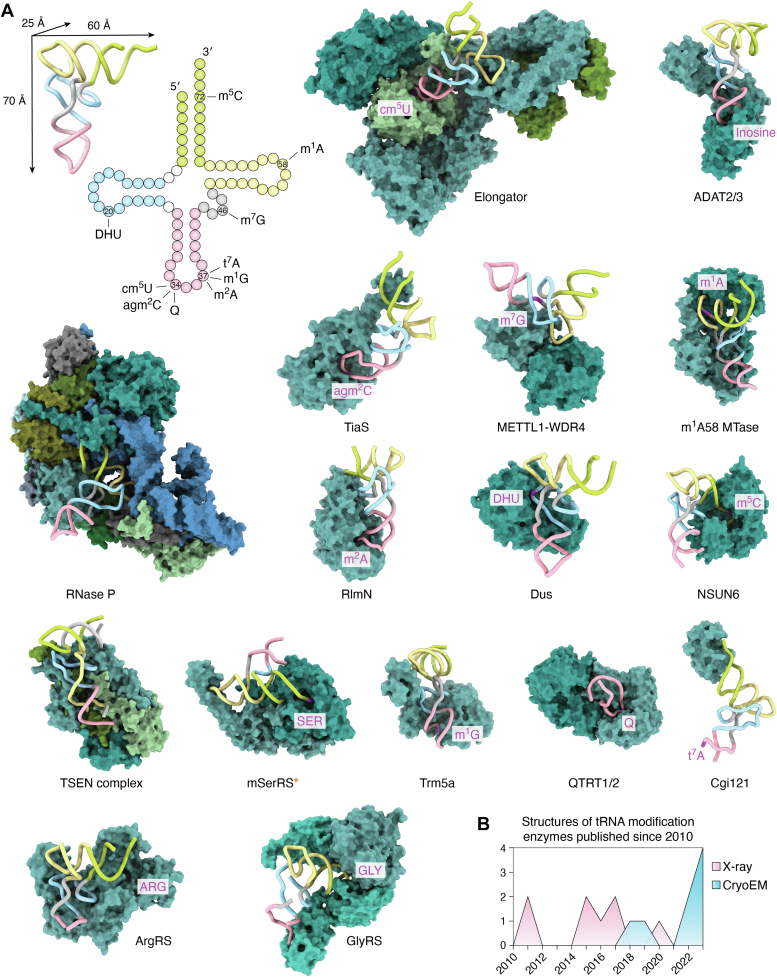
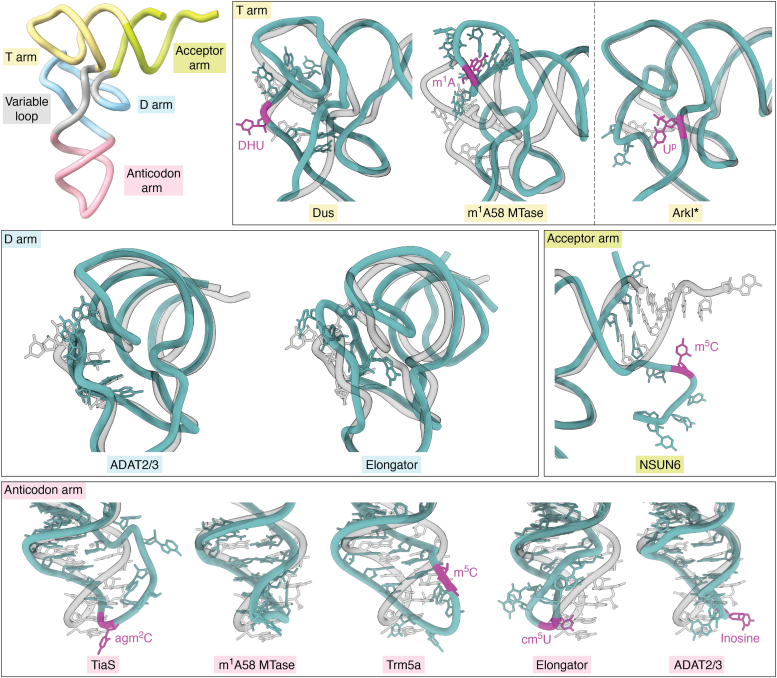
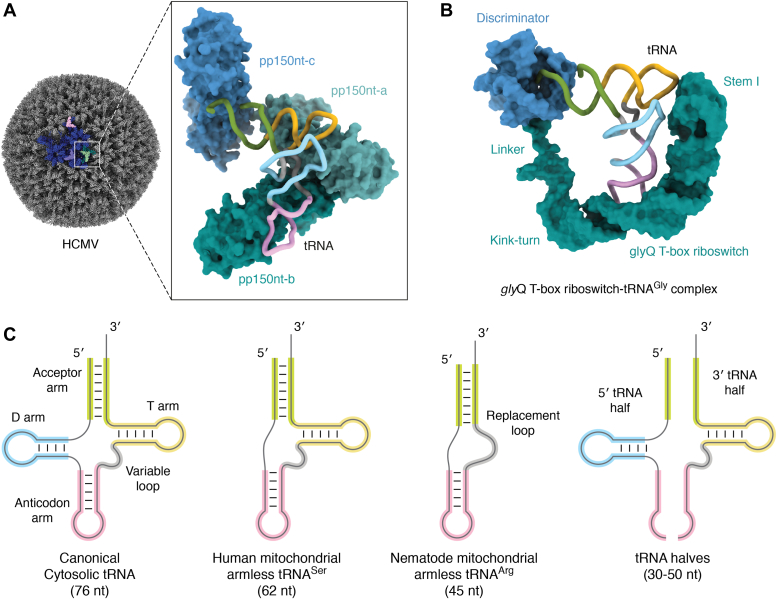
tRNAs are short noncoding RNAs responsible for decoding mRNA codon triplets, delivering correct amino acids to the ribosome, and mediating polypeptide chain formation. Due to their key roles during translation, tRNAs have a highly conserved shape and large sets of tRNAs are present in all living organisms. Regardless of sequence variability, all tRNAs fold into a relatively rigid three-dimensional L-shaped structure. The conserved tertiary organization of canonical tRNA arises through the formation of two orthogonal helices, consisting of the acceptor and anticodon domains. Both elements fold independently to stabilize the overall structure of tRNAs through intramolecular interactions between the D- and T-arm. During tRNA maturation, different modifying enzymes posttranscriptionally attach chemical groups to specific nucleotides, which not only affect translation elongation rates but also restrict local folding processes and confer local flexibility when required. The characteristic structural features of tRNAs are also employed by various maturation factors and modification enzymes to assure the selection, recognition, and positioning of specific sites within the substrate tRNAs. The cellular functional repertoire of tRNAs continues to extend well beyond their role in translation, partly, due to the expanding pool of tRNA-derived fragments. Here, we aim to summarize the most recent developments in the field to understand how three-dimensional structure affects the canonical and noncanonical functions of tRNA.
Keywords: NMR; RNA modifications; cryo-EM; crystallography; tRNA.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Crick F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958;12:138–163. - PubMed
-
- Roeder R.G., Rutter W.J. Multiple forms of DNA-dependent RNA polymerase in eukaryotic organisms. Nature. 1969;224:234–237. - PubMed
-
- Baranov P.V., Gesteland R.F., Atkins J.F. Recoding: translational bifurcations in gene expression. Gene. 2002;286:187–201. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
