

Reorientation of INO80 on hexasomes reveals basis for mechanistic versatility
- PMID: 37384669
- PMCID: PMC10480058
- DOI: 10.1126/science.adf4197
Reorientation of INO80 on hexasomes reveals basis for mechanistic versatility
Abstract
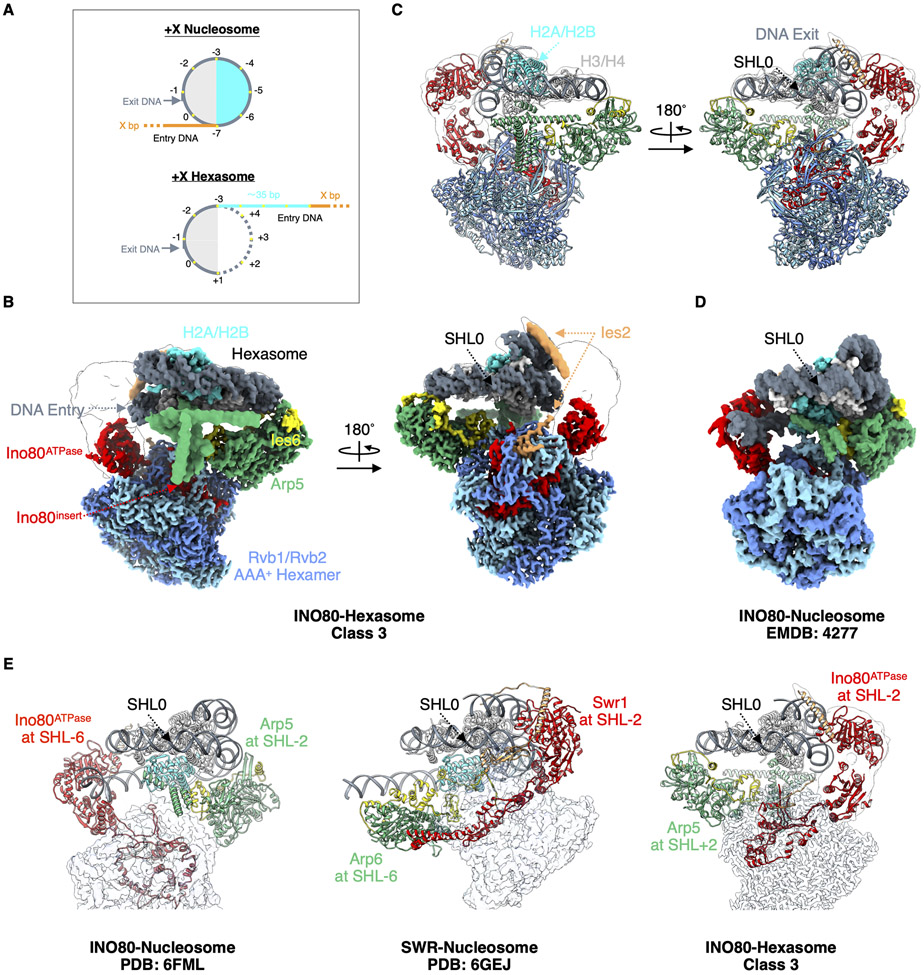
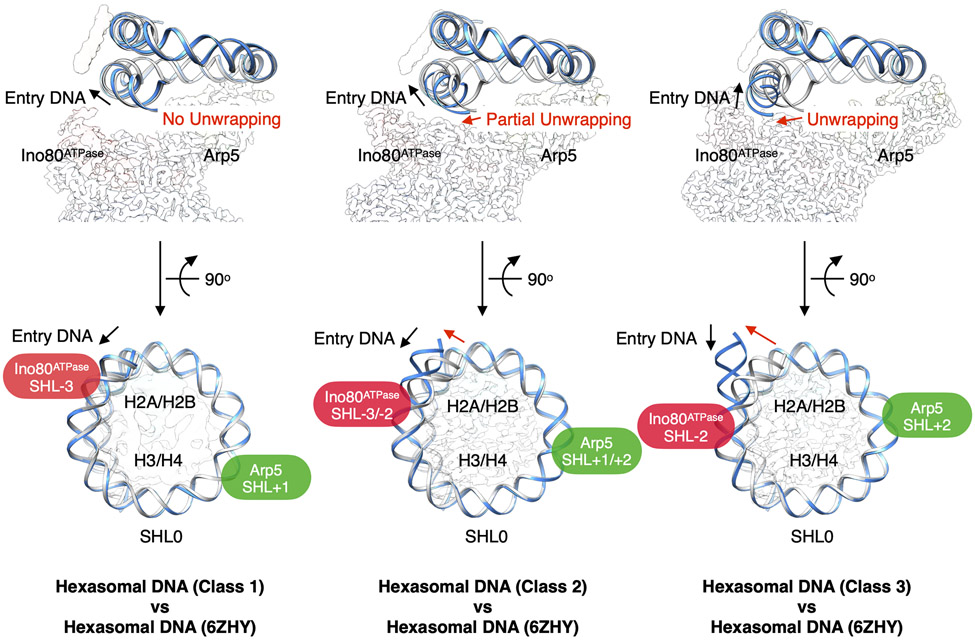
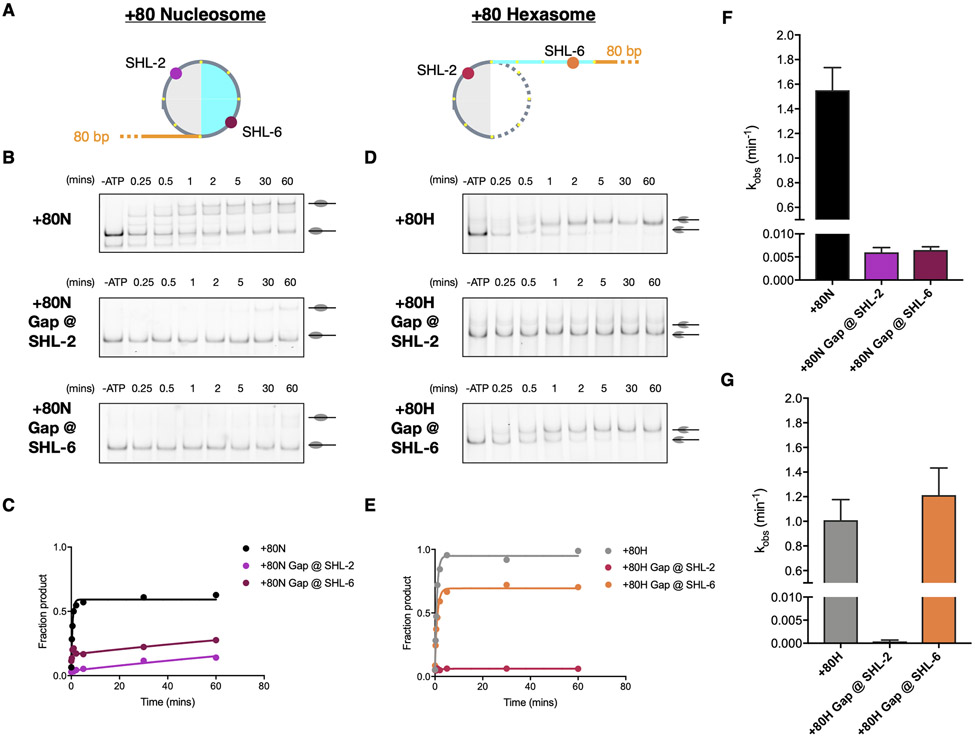
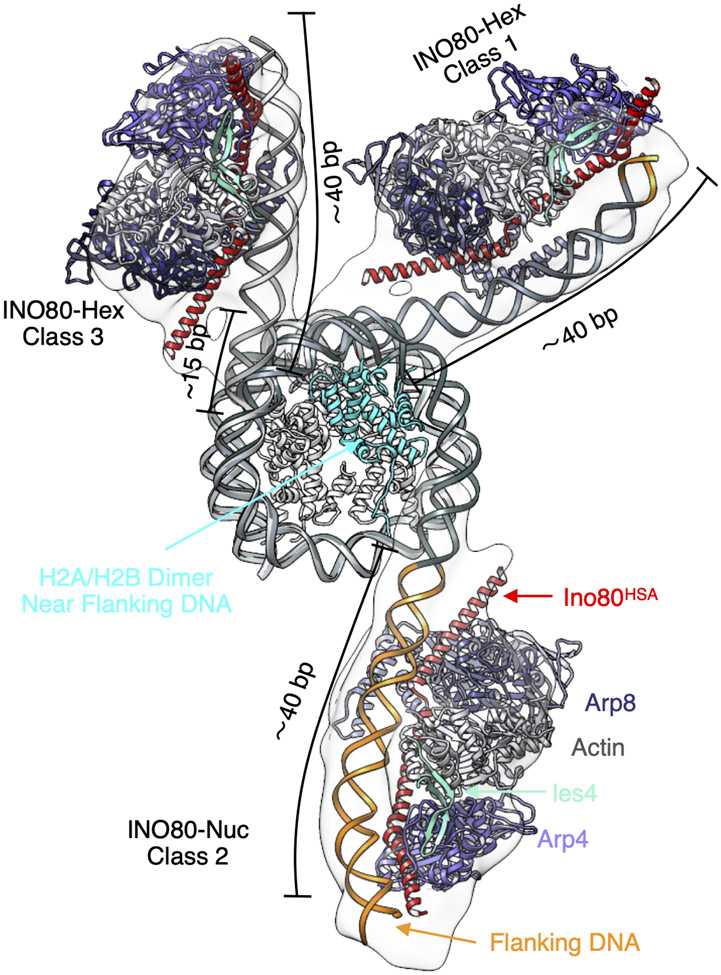
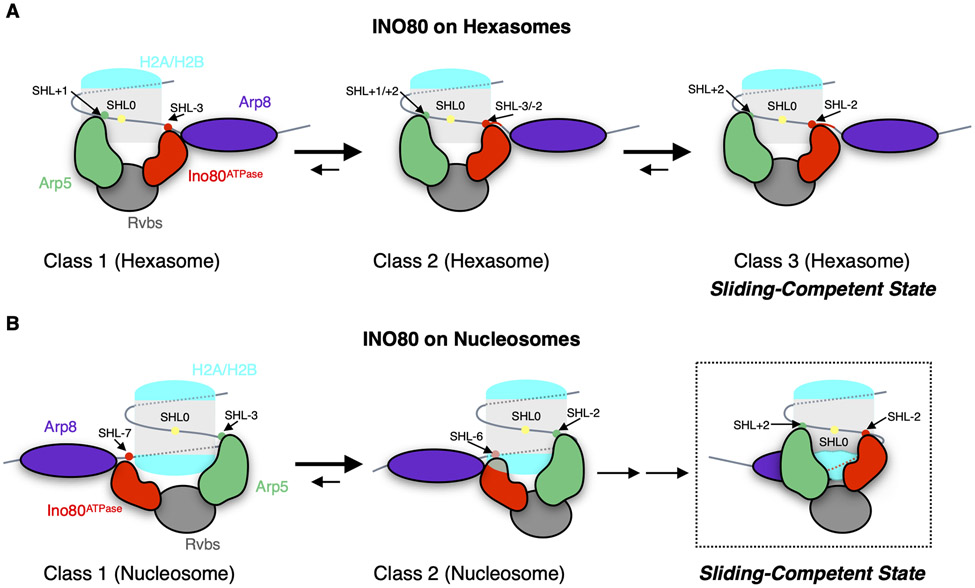
Unlike other chromatin remodelers, INO80 preferentially mobilizes hexasomes, which can form during transcription. Why INO80 prefers hexasomes over nucleosomes remains unclear. Here, we report structures of Saccharomyces cerevisiae INO80 bound to a hexasome or a nucleosome. INO80 binds the two substrates in substantially different orientations. On a hexasome, INO80 places its ATPase subunit, Ino80, at superhelical location -2 (SHL -2), in contrast to SHL -6 and SHL -7, as previously seen on nucleosomes. Our results suggest that INO80 action on hexasomes resembles action by other remodelers on nucleosomes such that Ino80 is maximally active near SHL -2. The SHL -2 position also plays a critical role for nucleosome remodeling by INO80. Overall, the mechanistic adaptations used by INO80 for preferential hexasome sliding imply that subnucleosomal particles play considerable regulatory roles.
Figures
Similar articles
-
A hexasome is the preferred substrate for the INO80 chromatin remodeling complex, allowing versatility of function.Mol Cell. 2022 Jun 2;82(11):2098-2112.e4. doi: 10.1016/j.molcel.2022.04.026. Epub 2022 May 20. Mol Cell. 2022. PMID: 35597239 Free PMC article.
-
To slide or not to slide: key role of the hexasome in chromatin remodeling revealed.Nat Struct Mol Biol. 2024 May;31(5):742-746. doi: 10.1038/s41594-024-01278-7. Epub 2024 May 20. Nat Struct Mol Biol. 2024. PMID: 38769465 Review.
-
Hexasome-INO80 complex reveals structural basis of noncanonical nucleosome remodeling.Science. 2023 Jul 21;381(6655):313-319. doi: 10.1126/science.adf6287. Epub 2023 Jun 29. Science. 2023. PMID: 37384673
-
Structural basis for ATP-dependent chromatin remodelling by the INO80 complex.Nature. 2018 Apr;556(7701):386-390. doi: 10.1038/s41586-018-0029-y. Epub 2018 Apr 11. Nature. 2018. PMID: 29643509 Free PMC article.
-
INO80 and SWR complexes: relating structure to function in chromatin remodeling.Trends Cell Biol. 2014 Nov;24(11):619-31. doi: 10.1016/j.tcb.2014.06.004. Epub 2014 Jul 31. Trends Cell Biol. 2014. PMID: 25088669 Review.
Cited by
-
In silico protein structural analysis of PRMT5 and RUVBL1 mutations arising in human cancers.Cancer Genet. 2025 Apr;292-293:49-56. doi: 10.1016/j.cancergen.2025.01.002. Epub 2025 Jan 17. Cancer Genet. 2025. PMID: 39874873 Free PMC article.
-
Conformational switching of Arp5 subunit differentially regulates INO80 chromatin remodeling.bioRxiv [Preprint]. 2024 May 11:2024.05.10.593625. doi: 10.1101/2024.05.10.593625. bioRxiv. 2024. Update in: Nucleic Acids Res. 2025 Jan 11;53(2):gkae1187. doi: 10.1093/nar/gkae1187. PMID: 38766108 Free PMC article. Updated. Preprint.
-
Conformational switching of Arp5 subunit regulates INO80 chromatin remodeling.Nucleic Acids Res. 2025 Jan 11;53(2):gkae1187. doi: 10.1093/nar/gkae1187. Nucleic Acids Res. 2025. PMID: 39676660 Free PMC article.
-
Structures of H2A.Z-associated human chromatin remodelers SRCAP and TIP60 reveal divergent mechanisms of chromatin engagement.bioRxiv [Preprint]. 2024 Jul 30:2024.07.30.605802. doi: 10.1101/2024.07.30.605802. bioRxiv. 2024. PMID: 39131301 Free PMC article. Preprint.
-
Hexasomal particles: consequence or also consequential?Curr Opin Genet Dev. 2024 Apr;85:102163. doi: 10.1016/j.gde.2024.102163. Epub 2024 Feb 26. Curr Opin Genet Dev. 2024. PMID: 38412564 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
