

Laminar specificity of the auditory perceptual awareness negativity: A biophysical modeling study
- PMID: 37384802
- PMCID: PMC10337981
- DOI: 10.1371/journal.pcbi.1011003
Laminar specificity of the auditory perceptual awareness negativity: A biophysical modeling study
Abstract
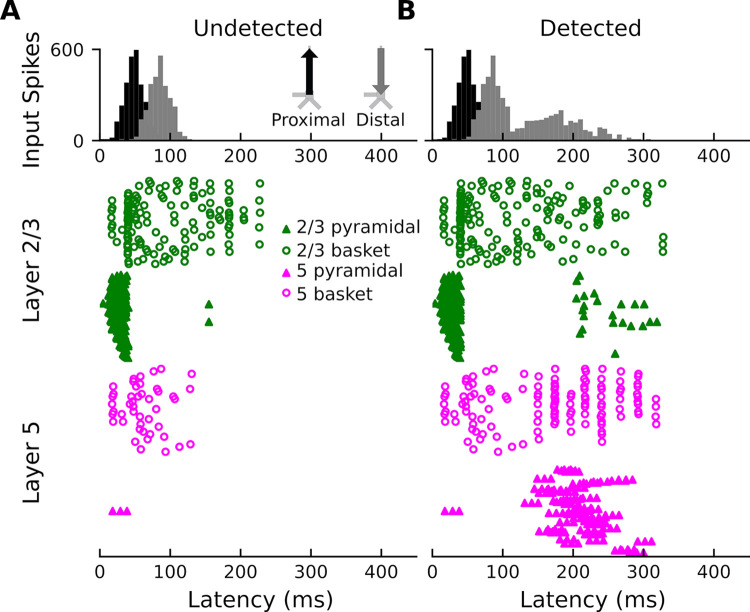
How perception of sensory stimuli emerges from brain activity is a fundamental question of neuroscience. To date, two disparate lines of research have examined this question. On one hand, human neuroimaging studies have helped us understand the large-scale brain dynamics of perception. On the other hand, work in animal models (mice, typically) has led to fundamental insight into the micro-scale neural circuits underlying perception. However, translating such fundamental insight from animal models to humans has been challenging. Here, using biophysical modeling, we show that the auditory awareness negativity (AAN), an evoked response associated with perception of target sounds in noise, can be accounted for by synaptic input to the supragranular layers of auditory cortex (AC) that is present when target sounds are heard but absent when they are missed. This additional input likely arises from cortico-cortical feedback and/or non-lemniscal thalamic projections and targets the apical dendrites of layer-5 (L5) pyramidal neurons. In turn, this leads to increased local field potential activity, increased spiking activity in L5 pyramidal neurons, and the AAN. The results are consistent with current cellular models of conscious processing and help bridge the gap between the macro and micro levels of perception-related brain activity.
Copyright: © 2023 Fernandez Pujol et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Update of
-
Laminar Specificity of the Auditory Perceptual Awareness Negativity: A Biophysical Modeling Study.bioRxiv [Preprint]. 2023 Mar 8:2023.03.06.531459. doi: 10.1101/2023.03.06.531459. bioRxiv. 2023. Update in: PLoS Comput Biol. 2023 Jun 29;19(6):e1011003. doi: 10.1371/journal.pcbi.1011003. PMID: 36945469 Free PMC article. Updated. Preprint.
References
-
- Crick F, Koch C. Toward a Neurobiological Theory of Consciousness. Seminars in the Neurosciences. 1990;2: 263–275.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
