

Germline loss-of-function PAM variants are enriched in subjects with pituitary hypersecretion
- PMID: 37388215
- PMCID: PMC10303134
- DOI: 10.3389/fendo.2023.1166076
Germline loss-of-function PAM variants are enriched in subjects with pituitary hypersecretion
Abstract
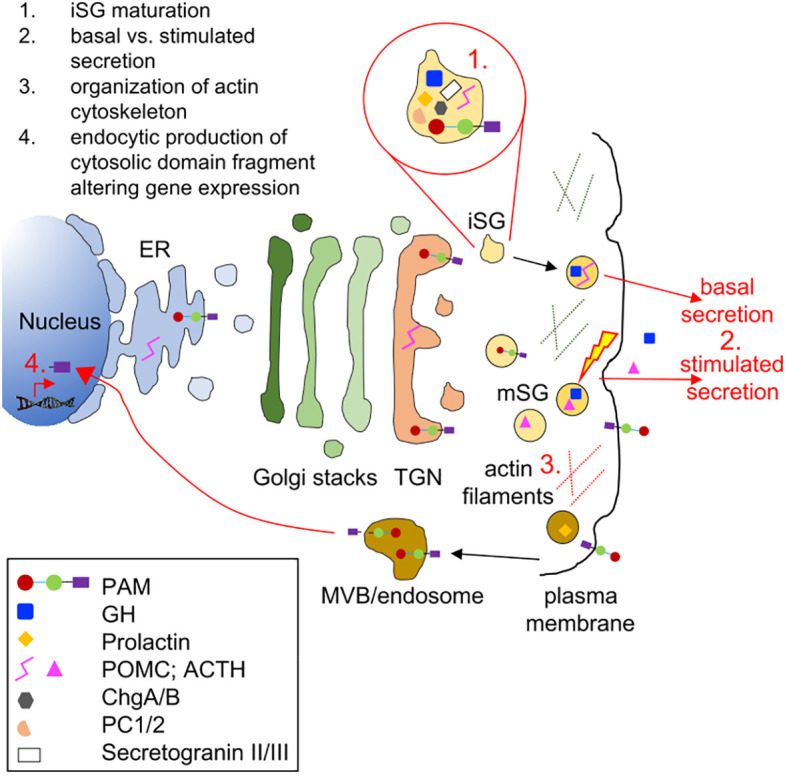
Introduction: Pituitary adenomas (PAs) are common, usually benign tumors of the anterior pituitary gland which, for the most part, have no known genetic cause. PAs are associated with major clinical effects due to hormonal dysregulation and tumoral impingement on vital brain structures. PAM encodes a multifunctional protein responsible for the essential C-terminal amidation of secreted peptides.
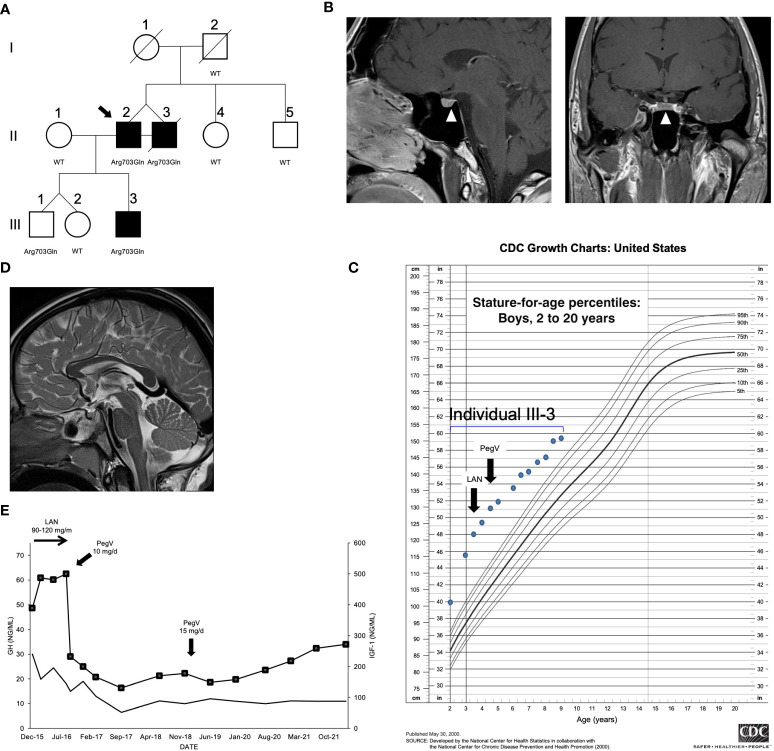
Methods: Following the identification of a loss-of-function variant (p.Arg703Gln) in the peptidylglycine a-amidating monooxygenase (PAM) gene in a family with pituitary gigantism, we investigated 299 individuals with sporadic PAs and 17 familial isolated PA kindreds for PAM variants. Genetic screening was performed by germline and tumor sequencing and germline copy number variation (CNV) analysis.
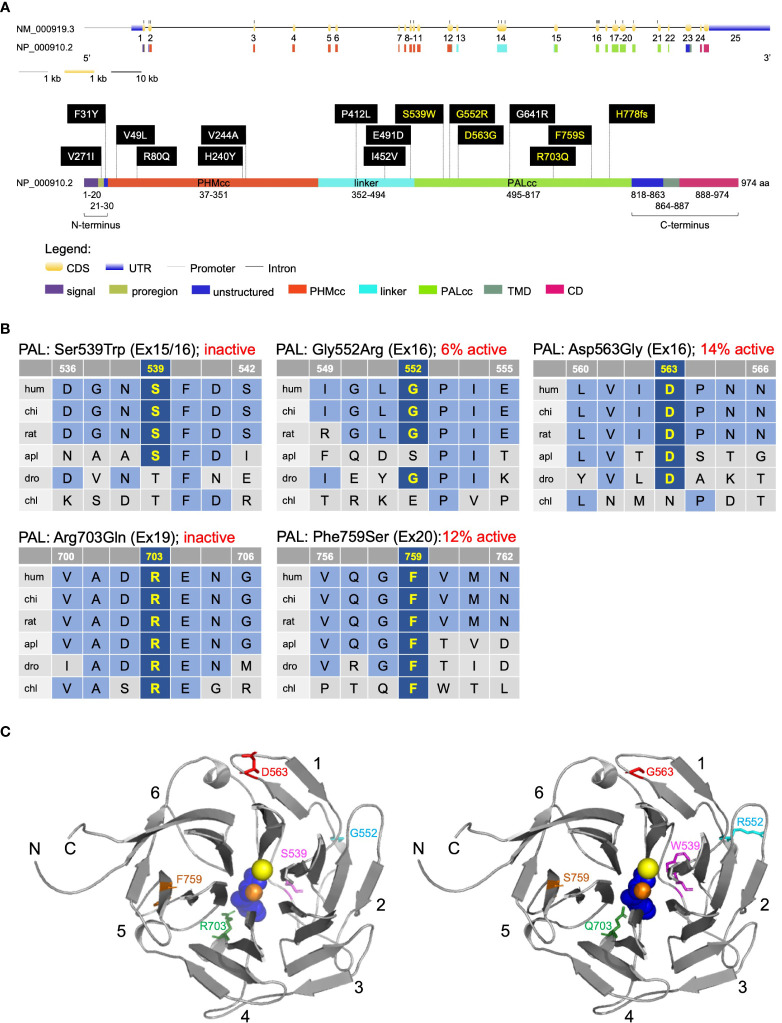
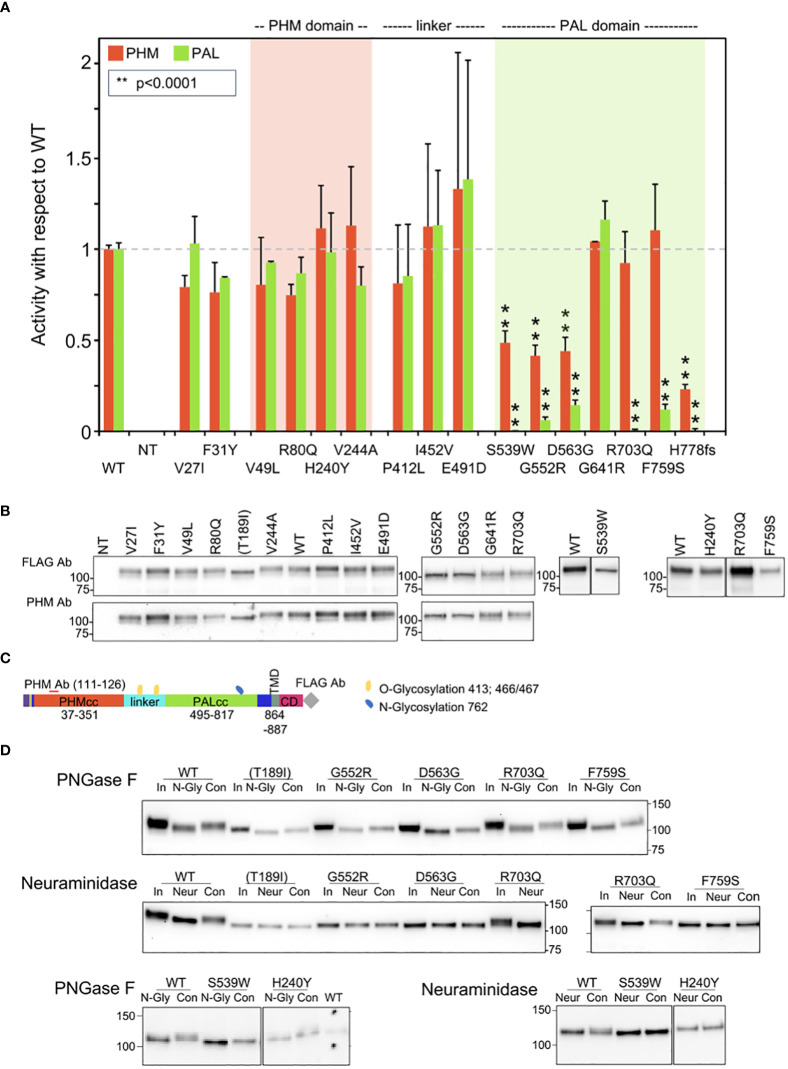
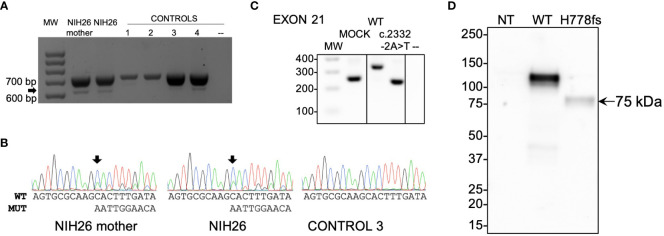
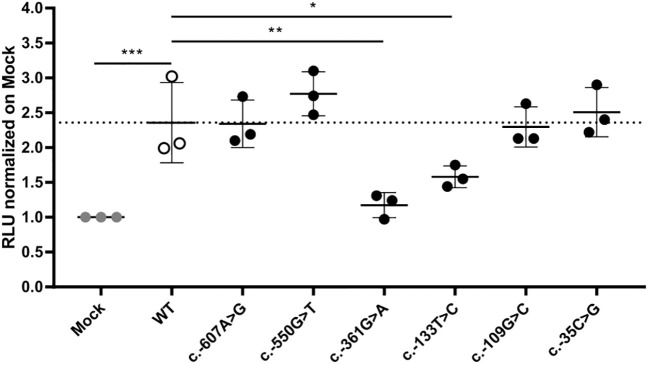
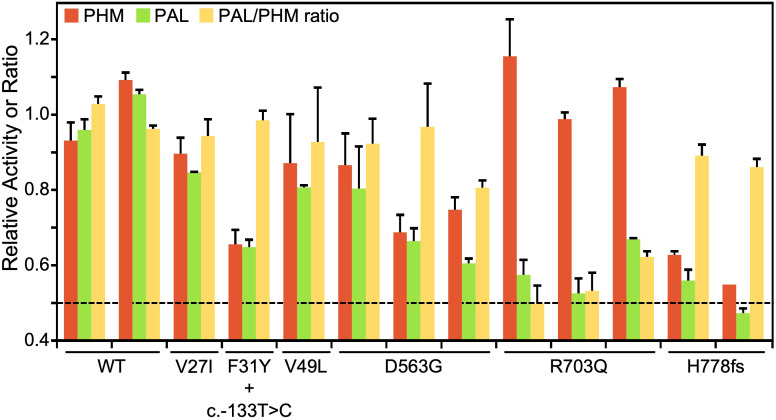
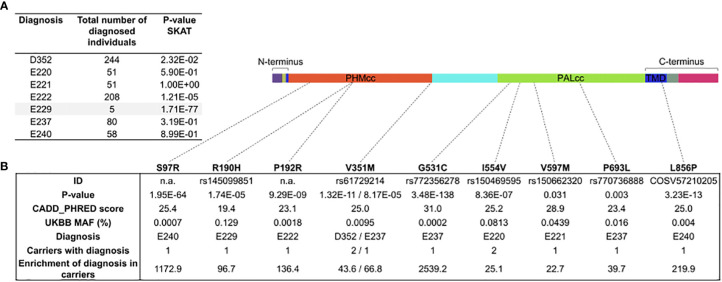
Results: In germline DNA, we detected seven heterozygous, likely pathogenic missense, truncating, and regulatory SNVs. These SNVs were found in sporadic subjects with growth hormone excess (p.Gly552Arg and p.Phe759Ser), pediatric Cushing disease (c.-133T>C and p.His778fs), or different types of PAs (c.-361G>A, p.Ser539Trp, and p.Asp563Gly). The SNVs were functionally tested in vitro for protein expression and trafficking by Western blotting, splicing by minigene assays, and amidation activity in cell lysates and serum samples. These analyses confirmed a deleterious effect on protein expression and/or function. By interrogating 200,000 exomes from the UK Biobank, we confirmed a significant association of the PAM gene and rare PAM SNVs with diagnoses linked to pituitary gland hyperfunction.
Conclusion: The identification of PAM as a candidate gene associated with pituitary hypersecretion opens the possibility of developing novel therapeutics based on altering PAM function.
Keywords: Cushing disease; acromegaly; amidation; gigantism; peptidylglycine α-amidating monooxygenase; pituitary tumors.
Copyright © 2023 Trivellin, Daly, Hernández-Ramírez, Araldi, Tatsi, Dale, Fridell, Mittal, Faucz, Iben, Li, Vitali, Stojilkovic, Kamenicky, Villa, Baussart, Chittiboina, Toro, Gahl, Eugster, Naves, Jaffrain-Rea, de Herder, Neggers, Petrossians, Beckers, Lania, Mains, Eipper and Stratakis.
Conflict of interest statement
AB, AD, FF, CS, and GT hold a patent on the GPR101 gene and its function US Patent No. 10,350,273, Treatment of Hormonal Disorders of Growth. CS holds patents on technologies involving PRKAR1A and related genes causing adrenal, pituitary, and other tumors. In addition, his laboratory has received research funding support by Pfizer Inc. for investigations on growth hormone–producing pituitary adenomas. CS has also consulted within the last 12 months with Lundbeck Pharmaceuticals and Sync, LLC, and is currently employed by ELPEN Pharmaceuticals. AB and AD have received research funding from Pfizer Inc. and Novo-Nordisk. M-LJ-R is part of the advisory board of Recordati Rare diseases since 2022. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Update of
-
Germline loss-of-function PAM variants are enriched in subjects with pituitary hypersecretion.medRxiv [Preprint]. 2023 Jan 20:2023.01.20.23284646. doi: 10.1101/2023.01.20.23284646. medRxiv. 2023. Update in: Front Endocrinol (Lausanne). 2023 Jun 14;14:1166076. doi: 10.3389/fendo.2023.1166076. PMID: 36711613 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous