

A multi-omics atlas of the human retina at single-cell resolution
- PMID: 37388908
- PMCID: PMC10300490
- DOI: 10.1016/j.xgen.2023.100298
A multi-omics atlas of the human retina at single-cell resolution
Abstract
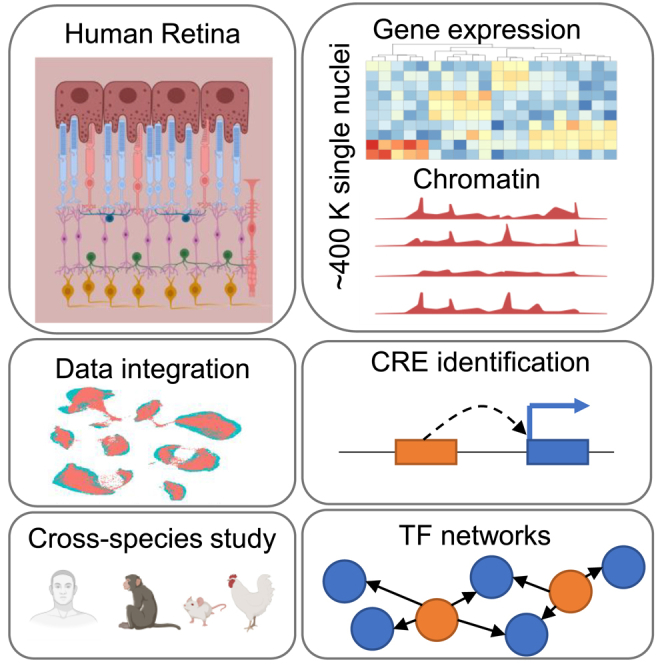
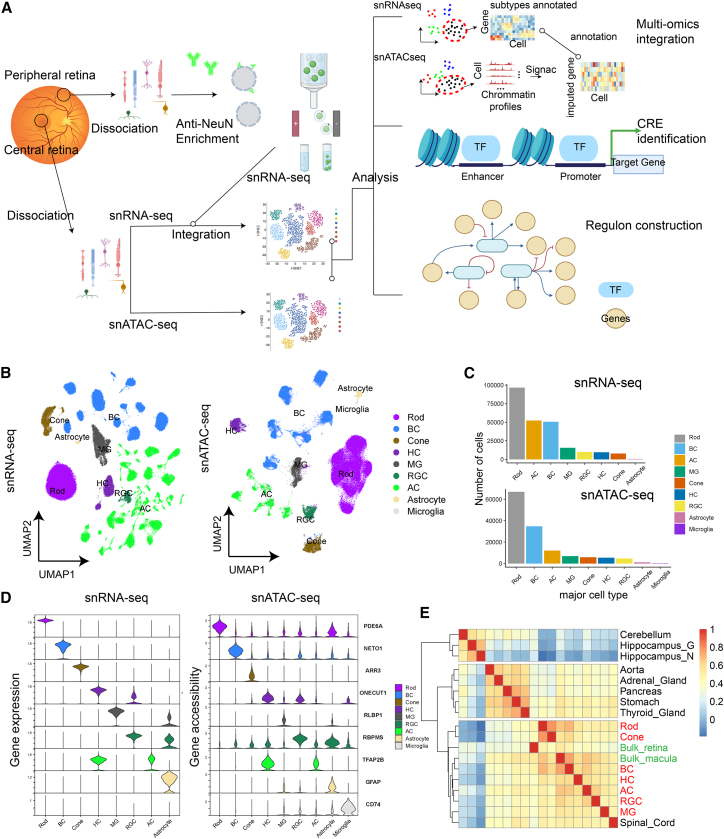
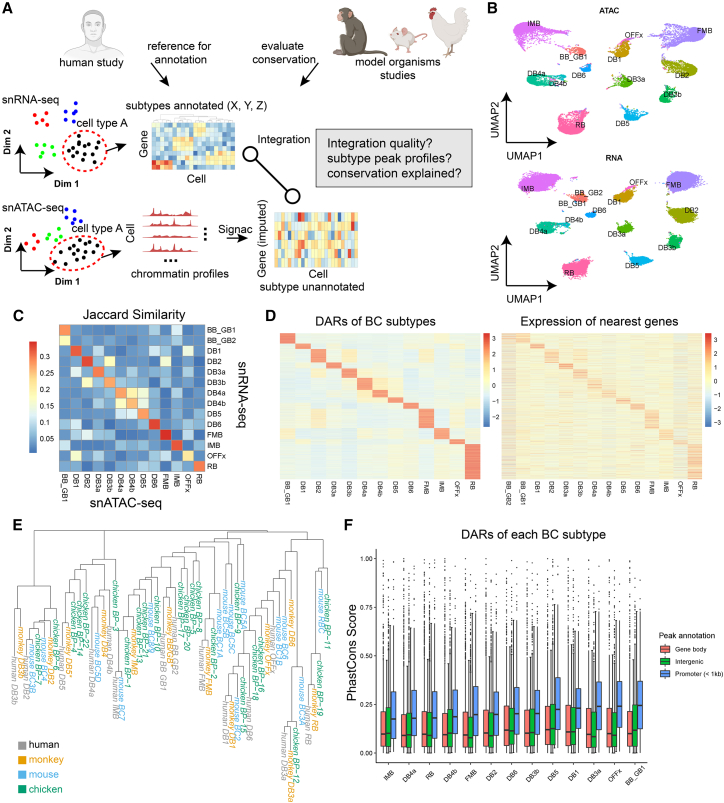
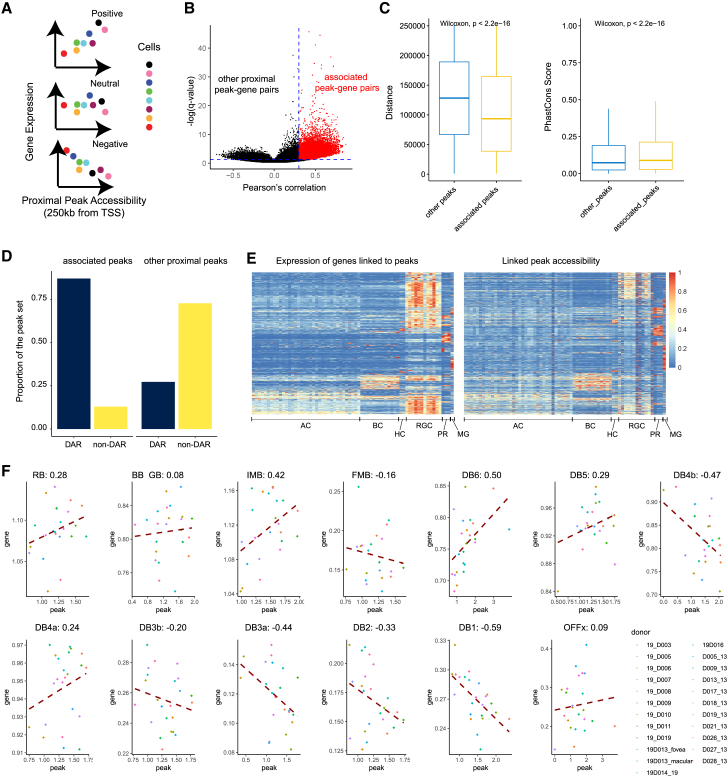
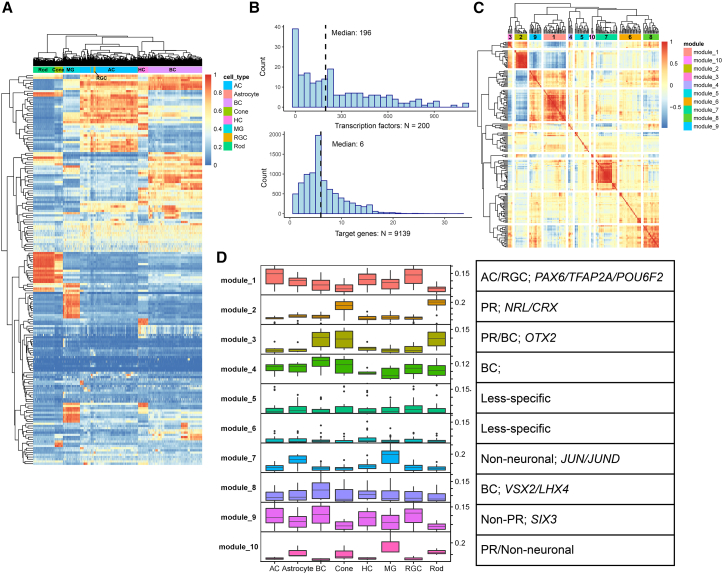
Cell classes in the human retina are highly heterogeneous with their abundance varying by several orders of magnitude. Here, we generated and integrated a multi-omics single-cell atlas of the adult human retina, including more than 250,000 nuclei for single-nuclei RNA-seq and 137,000 nuclei for single-nuclei ATAC-seq. Cross-species comparison of the retina atlas among human, monkey, mice, and chicken revealed relatively conserved and non-conserved types. Interestingly, the overall cell heterogeneity in primate retina decreases compared with that of rodent and chicken retina. Through integrative analysis, we identified 35,000 distal cis-element-gene pairs, constructed transcription factor (TF)-target regulons for more than 200 TFs, and partitioned the TFs into distinct co-active modules. We also revealed the heterogeneity of the cis-element-gene relationships in different cell types, even from the same class. Taken together, we present a comprehensive single-cell multi-omics atlas of the human retina as a resource that enables systematic molecular characterization at individual cell-type resolution.
Keywords: cross-species analysis; gene regulation; human retina; single-cell multi-omics.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Kolb H. Webvision; 2012. Gross anatomy of the eye.http://webvision.med.utah.edu/book/part-i-foundations/gross-anatomy-of-t... - PubMed
-
- Peng Y.R., Shekhar K., Yan W., Herrmann D., Sappington A., Bryman G.S., van Zyl T., Do M.T.H., Regev A., Sanes J.R. Molecular classification and comparative taxonomics of foveal and peripheral cells in primate retina. Cell. 2019;176:1222–1237.e22. doi: 10.1016/j.cell.2019.01.004. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
