

RNA-binding is an ancient trait of the Annexin family
- PMID: 37397259
- PMCID: PMC10311354
- DOI: 10.3389/fcell.2023.1161588
RNA-binding is an ancient trait of the Annexin family
Abstract
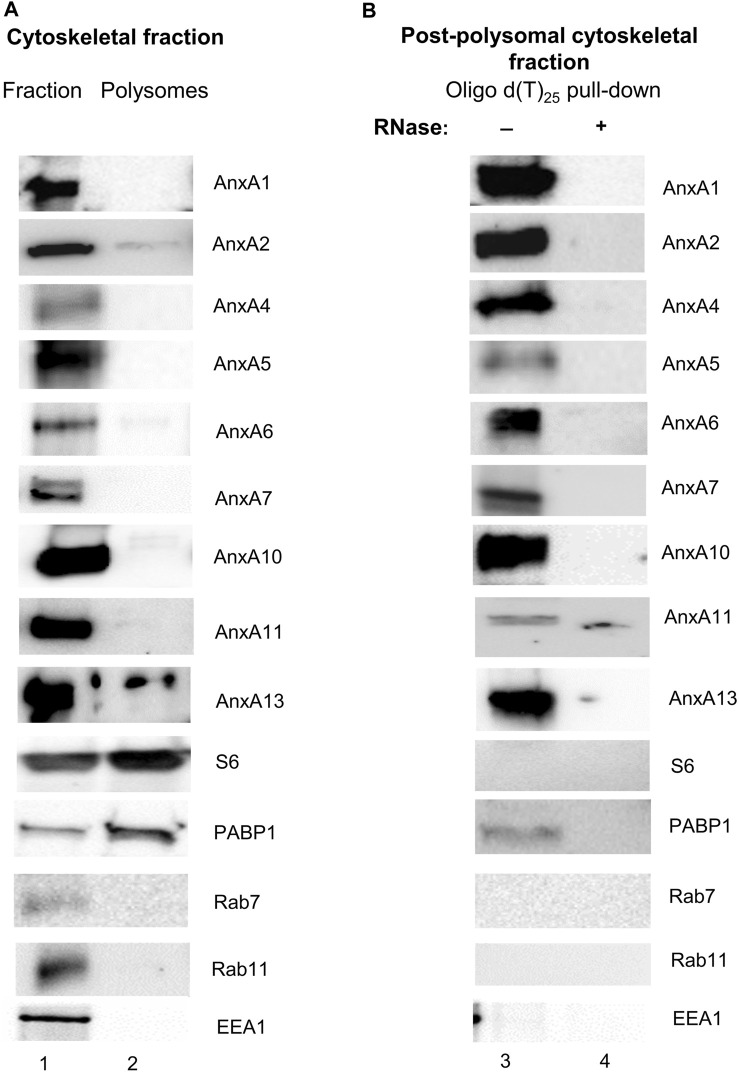
Introduction: The regulation of intracellular functions in mammalian cells involves close coordination of cellular processes. During recent years it has become evident that the sorting, trafficking and distribution of transport vesicles and mRNA granules/complexes are closely coordinated to ensure effective simultaneous handling of all components required for a specific function, thereby minimizing the use of cellular energy. Identification of proteins acting at the crossroads of such coordinated transport events will ultimately provide mechanistic details of the processes. Annexins are multifunctional proteins involved in a variety of cellular processes associated with Ca2+-regulation and lipid binding, linked to the operation of both the endocytic and exocytic pathways. Furthermore, certain Annexins have been implicated in the regulation of mRNA transport and translation. Since Annexin A2 binds specific mRNAs via its core structure and is also present in mRNP complexes, we speculated whether direct association with RNA could be a common property of the mammalian Annexin family sharing a highly similar core structure. Methods and results: Therefore, we performed spot blot and UV-crosslinking experiments to assess the mRNA binding abilities of the different Annexins, using annexin A2 and c-myc 3'UTRs as well as c-myc 5'UTR as baits. We supplemented the data with immunoblot detection of selected Annexins in mRNP complexes derived from the neuroendocrine rat PC12 cells. Furthermore, biolayer interferometry was used to determine the KD of selected Annexin-RNA interactions, which indicated distinct affinities. Amongst these Annexins, Annexin A13 and the core structures of Annexin A7, Annexin A11 bind c-myc 3'UTR with KDs in the nanomolar range. Of the selected Annexins, only Annexin A2 binds the c-myc 5'UTR indicating some selectivity. Discussion: The oldest members of the mammalian Annexin family share the ability to associate with RNA, suggesting that RNA-binding is an ancient trait of this protein family. Thus, the combined RNA- and lipid-binding properties of the Annexins make them attractive candidates to participate in coordinated long-distance transport of membrane vesicles and mRNAs regulated by Ca2+. The present screening results can thus pave the way for studies of the multifunctional Annexins in a novel cellular context.
Keywords: 3′untranslated region; Annexin; RNA granules; RNA-binding; c-Myc; mRNA; mRNP complexes.
Copyright © 2023 Patil, Panchal, Røstbø, Romanyuk, Hollås, Brenk, Grindheim and Vedeler.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Annexin A2 binds to the localization signal in the 3' untranslated region of c-myc mRNA.FEBS J. 2005 Jan;272(2):413-21. doi: 10.1111/j.1742-4658.2004.04481.x. FEBS J. 2005. PMID: 15654879
-
Comparative genetics and evolution of annexin A13 as the founder gene of vertebrate annexins.Mol Biol Evol. 2002 May;19(5):608-18. doi: 10.1093/oxfordjournals.molbev.a004120. Mol Biol Evol. 2002. PMID: 11961095
-
Annexins and Ca2+ handling in the heart.Cardiovasc Res. 2005 Mar 1;65(4):793-802. doi: 10.1016/j.cardiores.2004.11.010. Cardiovasc Res. 2005. PMID: 15721859 Review.
-
Annexin A2 binds the internal ribosomal entry site of c-myc mRNA and regulates its translation.RNA Biol. 2021 Oct 15;18(sup1):337-354. doi: 10.1080/15476286.2021.1947648. Epub 2021 Aug 4. RNA Biol. 2021. PMID: 34346292 Free PMC article.
-
S100-annexin complexes--structural insights.FEBS J. 2008 Oct;275(20):4956-66. doi: 10.1111/j.1742-4658.2008.06654.x. Epub 2008 Sep 13. FEBS J. 2008. PMID: 18795951 Review.
Cited by
-
Annexin, a Protein for All Seasons: From Calcium Dependent Membrane Metabolism to RNA Recognition.Bioessays. 2025 Jul;47(7):e70019. doi: 10.1002/bies.70019. Epub 2025 May 12. Bioessays. 2025. PMID: 40350993 Free PMC article. Review.
-
Annexins-a family of proteins with distinctive tastes for cell signaling and membrane dynamics.Nat Commun. 2024 Feb 21;15(1):1574. doi: 10.1038/s41467-024-45954-0. Nat Commun. 2024. PMID: 38383560 Free PMC article. Review.
-
Revisiting the role of Annexins in membrane trafficking.Cell Mol Life Sci. 2025 Jun 13;82(1):230. doi: 10.1007/s00018-025-05780-z. Cell Mol Life Sci. 2025. PMID: 40506619 Free PMC article. Review.
-
The structural properties of full-length annexin A11.Front Mol Biosci. 2024 Mar 7;11:1347741. doi: 10.3389/fmolb.2024.1347741. eCollection 2024. Front Mol Biosci. 2024. PMID: 38516187 Free PMC article.
References
-
- Anji A., Kumari M. (2011). A cis-acting region in the N-methyl-d-aspartate R1 3'-untranslated region interacts with the novel RNA-binding proteins beta subunit of alpha glucosidase II and annexin A2--effect of chronic ethanol exposure in vivo . Eur. J. Neurosci. 34 (8), 1200–1211. 10.1111/j.1460-9568.2011.07857.x - DOI - PMC - PubMed
-
- Aukrust I., Evensen L., Hollas H., Berven F., Atkinson R. A., Trave G., et al. (2006). Engineering, biophysical characterisation and binding properties of a soluble mutant form of annexin A2 domain IV that adopts a partially folded conformation. J. Mol. Biol. 363 (2), 469–481. 10.1016/j.jmb.2006.08.042 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous