

This is a preprint.
Rewiring capsule production by CRISPRi-based genetic oscillators demonstrates a functional role of phenotypic variation in pneumococcal-host interactions
- PMID: 37398107
- PMCID: PMC10312626
- DOI: 10.1101/2023.06.03.543575
Rewiring capsule production by CRISPRi-based genetic oscillators demonstrates a functional role of phenotypic variation in pneumococcal-host interactions
Update in
-
Synthetic genetic oscillators demonstrate the functional importance of phenotypic variation in pneumococcal-host interactions.Nat Commun. 2023 Nov 17;14(1):7454. doi: 10.1038/s41467-023-43241-y. Nat Commun. 2023. PMID: 37978173 Free PMC article.
Abstract
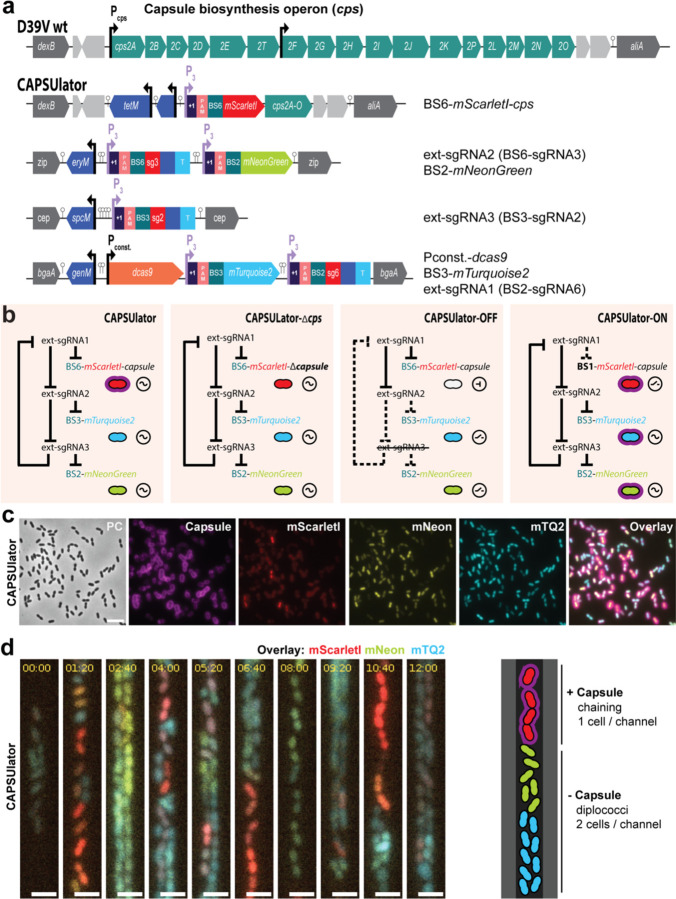
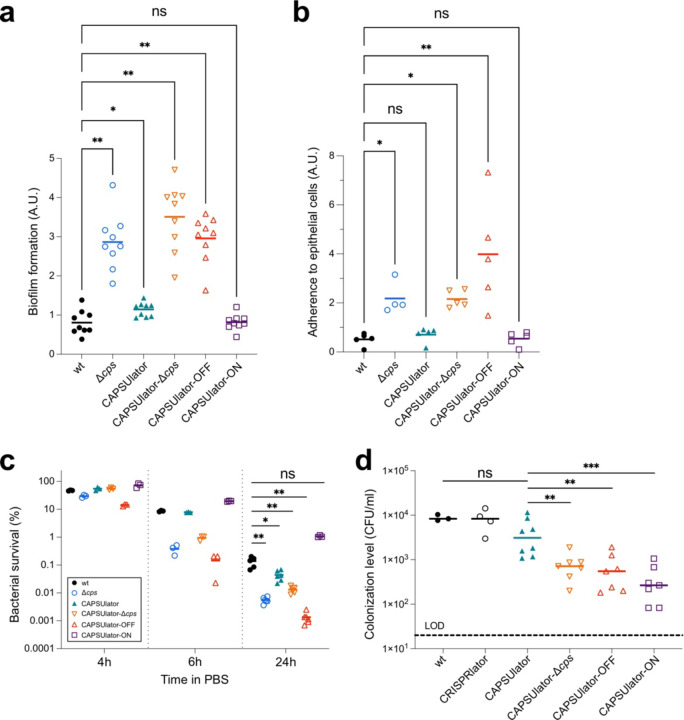
Phenotypic variation is the phenomenon in which clonal cells display different traits even under identical environmental conditions. This plasticity is thought to be important for processes including bacterial virulence1-8, but direct evidence for its relevance is often lacking. For instance, variation in capsule production in the human pathogen Streptococcus pneumoniae has been linked to different clinical outcomes9-14, but the exact relationship between variation and pathogenesis is not well understood due to complex natural regulation15-20. In this study, we used synthetic oscillatory gene regulatory networks (GRNs) based on CRISPR interference together with live cell microscopy and cell tracking within microfluidics devices to mimic and test the biological function of bacterial phenotypic variation. We provide a universally applicable approach for engineering intricate GRNs using only two components: dCas9 and extended sgRNAs (ext-sgRNAs). Our findings demonstrate that variation in capsule production is beneficial for pneumococcal fitness in traits associated with pathogenesis providing conclusive evidence for this longstanding question.
Conflict of interest statement
Competing interests Authors declare no competing interests.
Figures
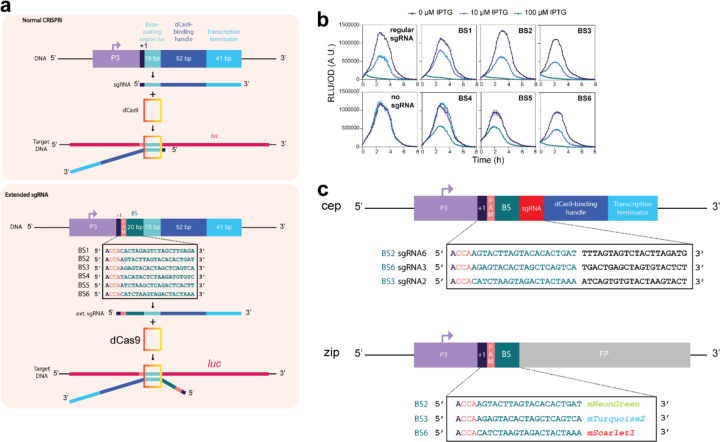
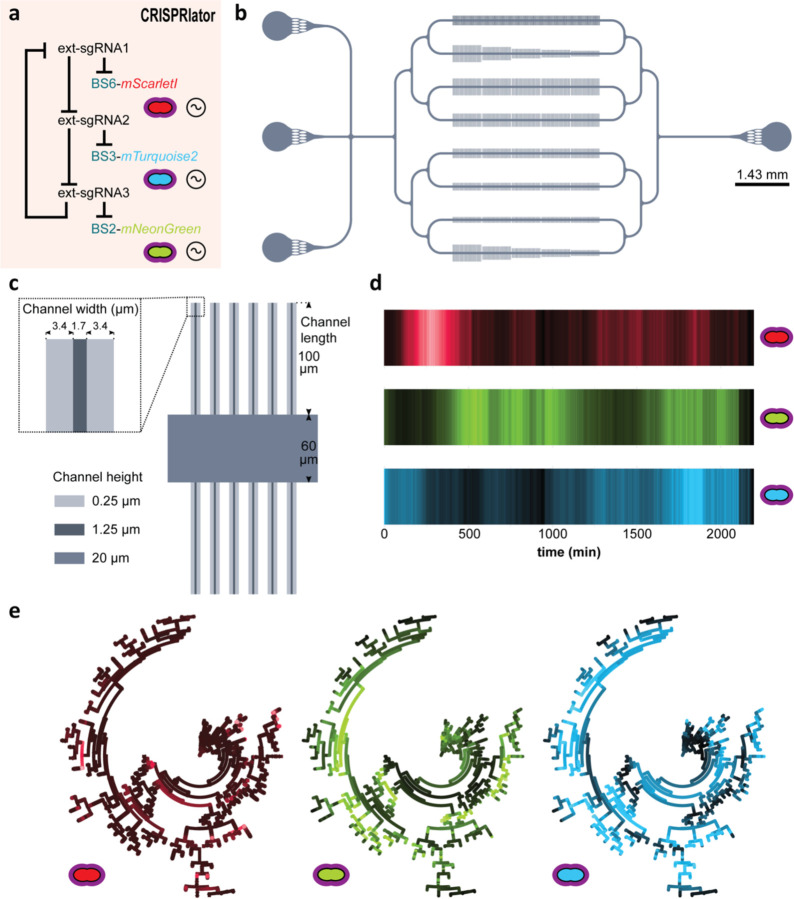
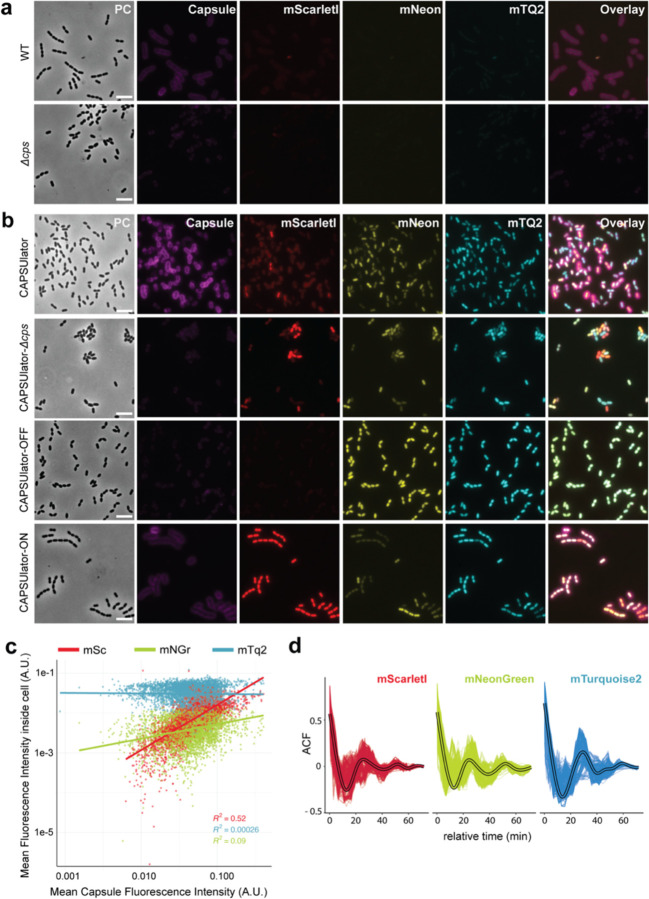
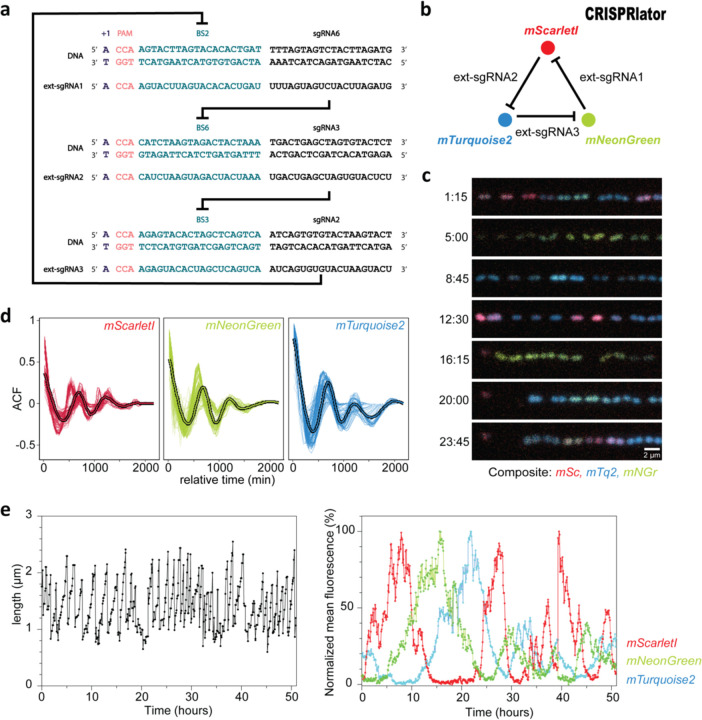
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources