

This is a preprint.
Guanine-containing ssDNA and RNA induce dimeric and tetrameric SAMHD1 in cryo-EM and binding studies
- PMID: 37398126
- PMCID: PMC10312740
- DOI: 10.1101/2023.06.15.544806
Guanine-containing ssDNA and RNA induce dimeric and tetrameric SAMHD1 in cryo-EM and binding studies
Update in
-
Guanine-containing ssDNA and RNA induce dimeric and tetrameric structural forms of SAMHD1.Nucleic Acids Res. 2023 Dec 11;51(22):12443-12458. doi: 10.1093/nar/gkad971. Nucleic Acids Res. 2023. PMID: 37930833 Free PMC article.
Abstract
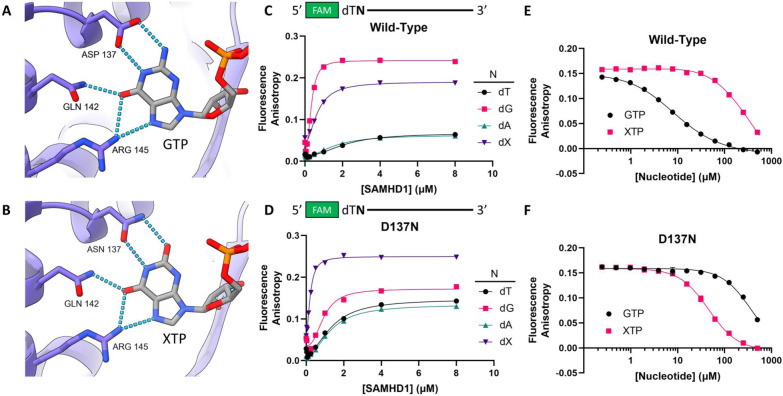
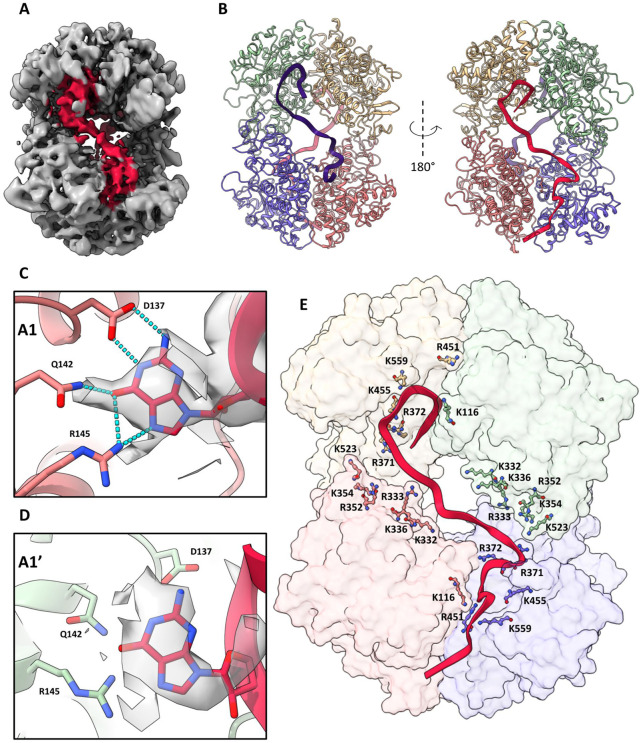
The dNTPase activity of tetrameric SAM and HD domain containing deoxynucleoside triphosphate triphosphohydrolase 1 (SAMHD1) plays a critical role in cellular dNTP regulation. SAMHD1 also associates with stalled DNA replication forks, DNA repair foci, ssRNA, and telomeres. The above functions require nucleic acid binding by SAMHD1, which may be modulated by its oligomeric state. Here we establish that the guanine-specific A1 activator site of each SAMHD1 monomer is used to target the enzyme to guanine nucleotides within single-stranded (ss) DNA and RNA. Remarkably, nucleic acid strands containing a single guanine base induce dimeric SAMHD1, while two or more guanines with ~20 nucleotide spacing induce a tetrameric form. A cryo-EM structure of ssRNA-bound tetrameric SAMHD1 shows how ssRNA strands bridge two SAMHD1 dimers and stabilize the structure. This ssRNA-bound tetramer is inactive with respect to dNTPase and RNase activity.
Conflict of interest statement
Conflict of interest statement M.W.S, Y.X., K.W.H, S.H., B.B., and D.S. are employees of Pfizer Inc. and may hold shares in the company.
Figures





References
-
- Zhang Z. et al. Involvement of SAMHD1 in dNTP homeostasis and the maintenance of genomic integrity and oncotherapy (Review). Int. J. Oncol. 56, 879–888 (2020). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous