

This is a preprint.
The basis of antigenic operon fragmentation in Bacteroidota and commensalism
- PMID: 37398285
- PMCID: PMC10312583
- DOI: 10.1101/2023.06.02.543472
The basis of antigenic operon fragmentation in Bacteroidota and commensalism
Update in
-
Antigenic operon fragmentation and diversification mechanism in Bacteroidota impacts gut metagenomics and pathobionts in Crohn's disease microlesions.Gut Microbes. 2024 Jan-Dec;16(1):2350150. doi: 10.1080/19490976.2024.2350150. Epub 2024 Jun 6. Gut Microbes. 2024. PMID: 38841888 Free PMC article.
Abstract
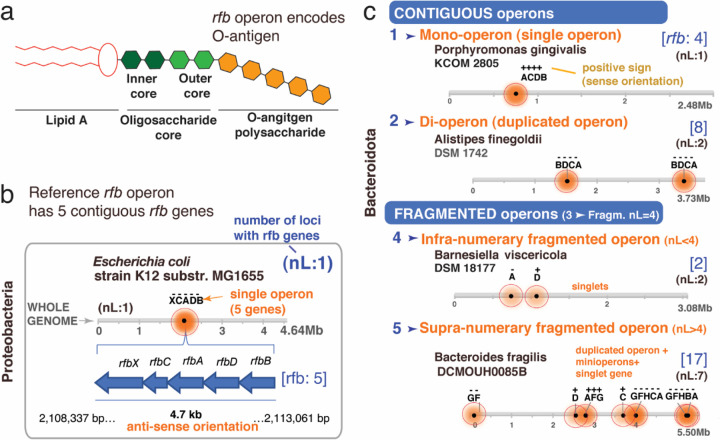
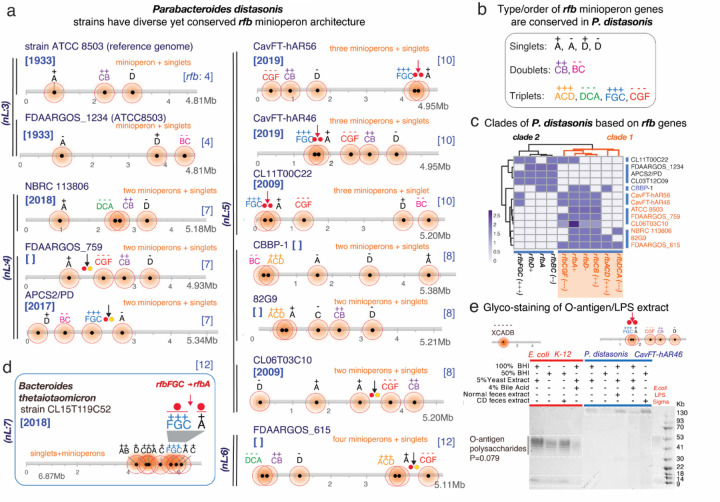
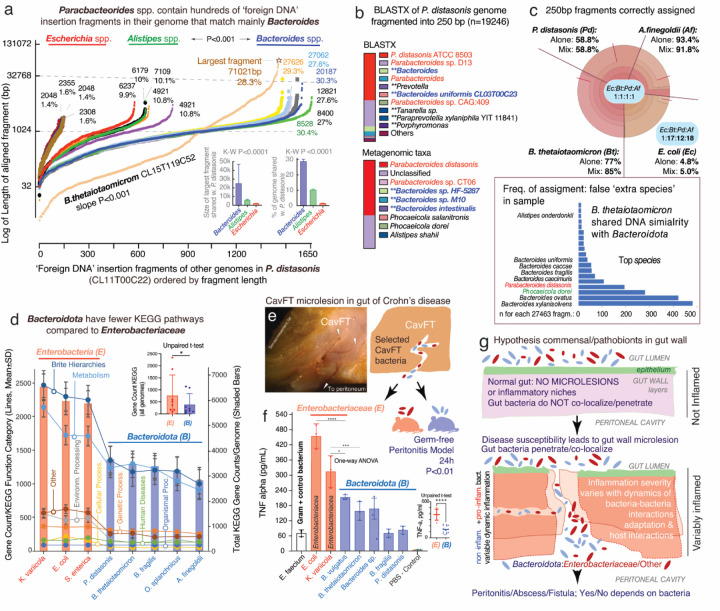
The causes for variability of pro-inflammatory surface antigens that affect gut commensal/opportunistic dualism within the phylum Bacteroidota remain unclear (1, 2). Using the classical lipopolysaccharide/O-antigen 'rfb operon' in Enterobacteriaceae as a surface antigen model (5-gene-cluster rfbABCDX), and a recent rfbA-typing strategy for strain classification (3), we characterized the architecture/conservancy of the entire rfb operon in Bacteroidota. Analyzing complete genomes, we discovered that most Bacteroidota have the rfb operon fragmented into non-random gene-singlets and/or doublets/triplets, termed 'minioperons'. To reflect global operon integrity, duplication, and fragmentation principles, we propose a five-category (infra/supernumerary) cataloguing system and a Global Operon Profiling System for bacteria. Mechanistically, genomic sequence analyses revealed that operon fragmentation is driven by intra-operon insertions of predominantly Bacteroides-DNA (thetaiotaomicron/fragilis) and likely natural selection in specific micro-niches. Bacteroides-insertions, also detected in other antigenic operons (fimbriae), but not in operons deemed essential (ribosomal), could explain why Bacteroidota have fewer KEGG-pathways despite large genomes (4). DNA insertions overrepresenting DNA-exchange-avid species, impact functional metagenomics by inflating gene-based pathway inference and overestimating 'extra-species' abundance. Using bacteria from inflammatory gut-wall cavernous micro-tracts (CavFT) in Crohn's Disease (5), we illustrate that bacteria with supernumerary-fragmented operons cannot produce O-antigen, and that commensal/CavFT Bacteroidota stimulate macrophages with lower potency than Enterobacteriaceae, and do not induce peritonitis in mice. The impact of 'foreign-DNA' insertions on pro-inflammatory operons, metagenomics, and commensalism offers potential for novel diagnostics and therapeutics.
Keywords: Alistipes; Bacteroides; Escherichia coli; O-antigen; Parabacteroides; Prevotella; Salmonella.
Conflict of interest statement
Conflict of Interest: The authors declare they have no conflicts of interest.
Figures



References
-
- Bank NC, Singh V, Rodriguez-Palacios A. Classification of Parabacteroides distasonis and other Bacteroidetes using O-antigen virulence gene: RfbA-Typing and hypothesis for pathogenic vs. probiotic strain differentiation. Gut Microbes. 2022;14(1):1997293. Epub 2022/01/30. doi: 10.1080/19490976.2021.1997293. - DOI - PMC - PubMed
-
- Yang F, Kumar A, Davenport KW, Kelliher JM, Ezeji JC, Good CE, et al. Complete Genome Sequence of a Parabacteroides distasonis Strain (CavFT hAR46) Isolated from a Gut Wall-Cavitating Microlesion in a Patient with Severe Crohn’s Disease. Microbiol Resour Announc. 2019;8(36). Epub 2019/09/05. doi: 10.1128/MRA.00585-19. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources