

The evolution of archaeal flagellar filaments
- PMID: 37399404
- PMCID: PMC10334743
- DOI: 10.1073/pnas.2304256120
The evolution of archaeal flagellar filaments
Abstract
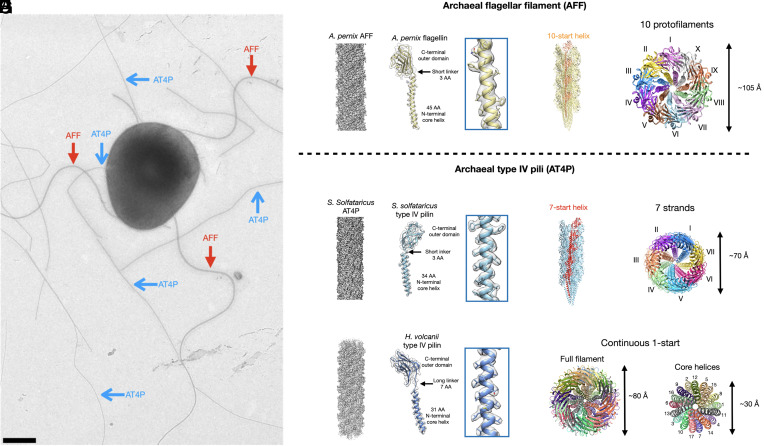
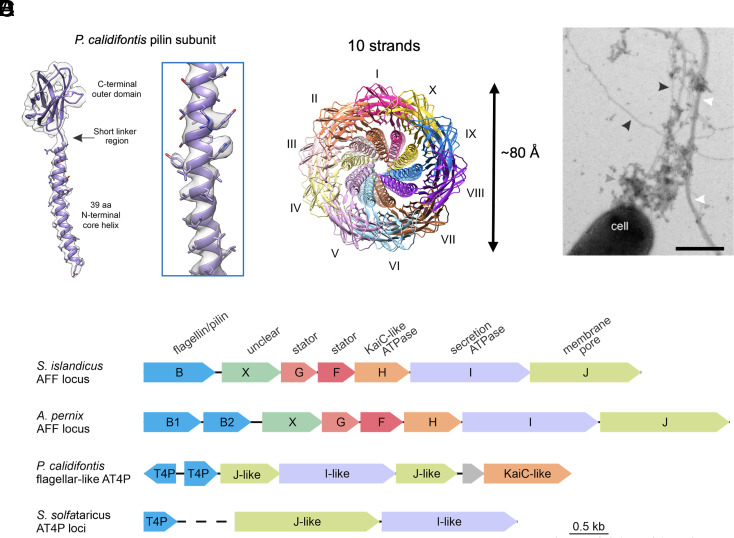
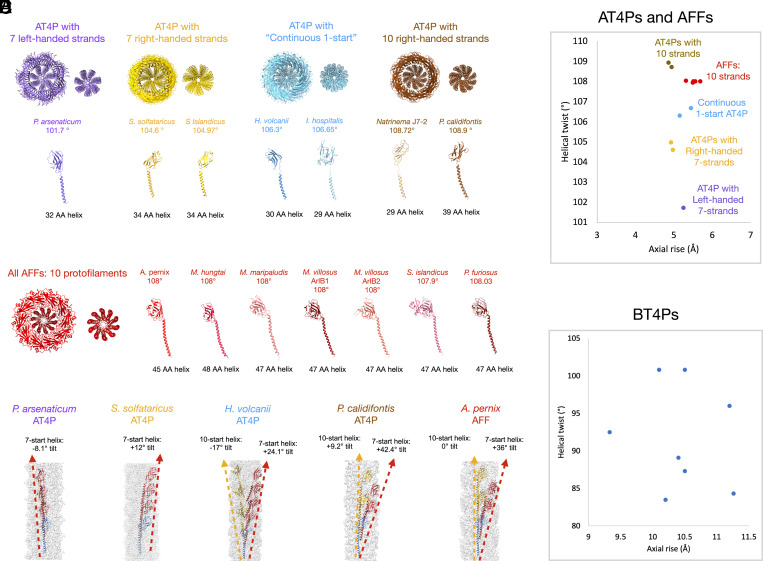
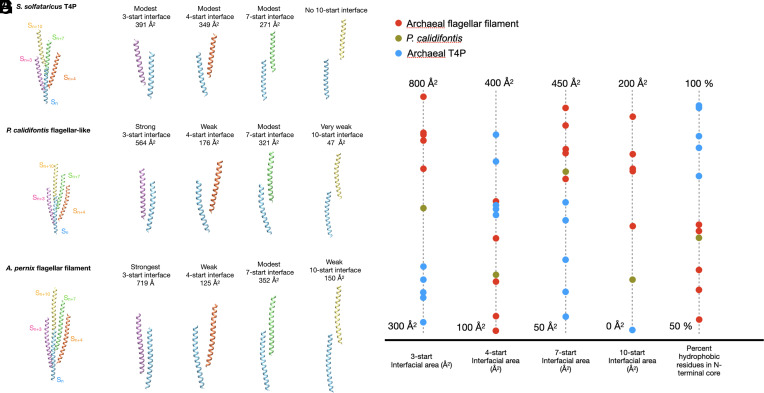
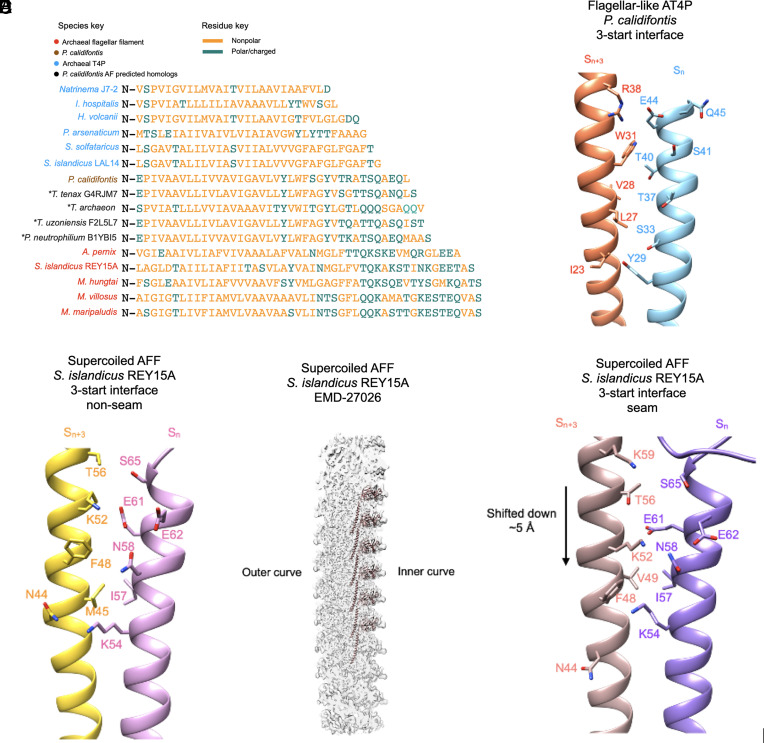
Flagellar motility has independently arisen three times during evolution: in bacteria, archaea, and eukaryotes. In prokaryotes, the supercoiled flagellar filaments are composed largely of a single protein, bacterial or archaeal flagellin, although these two proteins are not homologous, while in eukaryotes, the flagellum contains hundreds of proteins. Archaeal flagellin and archaeal type IV pilin are homologous, but how archaeal flagellar filaments (AFFs) and archaeal type IV pili (AT4Ps) diverged is not understood, in part, due to the paucity of structures for AFFs and AT4Ps. Despite having similar structures, AFFs supercoil, while AT4Ps do not, and supercoiling is essential for the function of AFFs. We used cryo-electron microscopy to determine the atomic structure of two additional AT4Ps and reanalyzed previous structures. We find that all AFFs have a prominent 10-strand packing, while AT4Ps show a striking structural diversity in their subunit packing. A clear distinction between all AFF and all AT4P structures involves the extension of the N-terminal α-helix with polar residues in the AFFs. Additionally, we characterize a flagellar-like AT4P from Pyrobaculum calidifontis with filament and subunit structure similar to that of AFFs which can be viewed as an evolutionary link, showing how the structural diversity of AT4Ps likely allowed for an AT4P to evolve into a supercoiling AFF.
Keywords: cryo-EM; helical polymers; motility.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Alam M., Oesterhelt D., Morphology, function and isolation of halobacterial flagella. J. Mol. Biol. 176, 459–475 (1984). - PubMed
-
- Brown H. P., On the structure and mechanics of the protozoan flagellum. Ohio J. Sci. 45, 247–301 (1945). - PubMed
-
- Pijper A., Abraham G., Wavelengths of bacterial flagella. J. Gen. Microbiol. 10, 452–456 (1954). - PubMed
-
- Hyman H. C., Trachtenberg S., Point mutations that lock Salmonella typhimurium flagellar filaments in the straight right-handed and left-handed forms and their relation to filament superhelicity. J. Mol. Biol. 220, 79–88 (1991). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
