

A ribonuclease activity linked to DYW1 in vitro is inhibited by RIP/MORF proteins
- PMID: 37400527
- PMCID: PMC10318007
- DOI: 10.1038/s41598-023-36969-6
A ribonuclease activity linked to DYW1 in vitro is inhibited by RIP/MORF proteins
Abstract
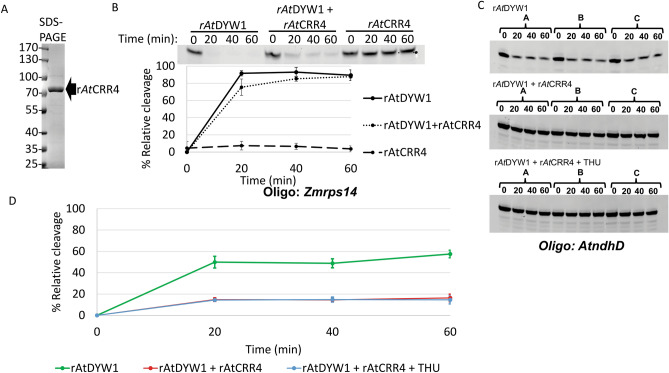
Organellar C-to-U RNA editing in plants occurs in complexes composed of various classes of nuclear-encoded proteins. DYW-deaminases are zinc metalloenzymes that catalyze hydrolytic deamination required for C-to-U modification editing. Solved crystal structures for DYW-deaminase domains display all structural features consistent with a canonical cytidine deamination mechanism. However, some recombinant DYW-deaminases from plants have been associated with ribonuclease activity in vitro. Direct ribonuclease activity by an editing factor is confounding since it is not required for deamination of cytosine, theoretically would be inimical for mRNA editing, and does not have a clear physiological function in vivo. His-tagged recombinant DYW1 from Arabidopsis thaliana (rAtDYW1) was expressed and purified using immobilized metal affinity chromatography (IMAC). Fluorescently labeled RNA oligonucleotides were incubated with recombinant AtDYW1 under different conditions. Percent relative cleavage of RNA probes was recorded at multiple time points from triplicate reactions. The effects of treatment with zinc chelators EDTA and 1, 10-phenanthroline were examined for rAtDYW1. Recombinant His-tagged RNA editing factors AtRIP2, ZmRIP9, AtRIP9, AtOZ1, AtCRR4, and AtORRM1 were expressed in E. coli and purified. Ribonuclease activity was assayed for rAtDYW1 in the presence of different editing factors. Lastly, the effects on nuclease activity in the presence of nucleotides and modified nucleosides were investigated. RNA cleavage observed in this study was linked to the recombinant editing factor rAtDYW1 in vitro. The cleavage reaction is sensitive to high concentrations of zinc chelators, indicating a role for zinc ions for activity. The addition of equal molar concentrations of recombinant RIP/MORF proteins reduced cleavage activity associated with rAtDYW1. However, addition of equal molar concentrations of purified recombinant editing complex proteins AtCRR4, AtORRM1, and AtOZ1 did not strongly inhibit ribonuclease activity on RNAs lacking an AtCRR4 cis-element. Though AtCRR4 inhibited AtDYW1 activity for oligonucleotides with a cognate cis-element. The observation that editing factors limit ribonuclease activity of rAtDYW1 in vitro, suggests that nuclease activities are limited to RNAs in absence of native editing complex partners. Purified rAtDYW1 was associated with the hydrolysis of RNA in vitro, and activity was specifically inhibited by RNA editing factors.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

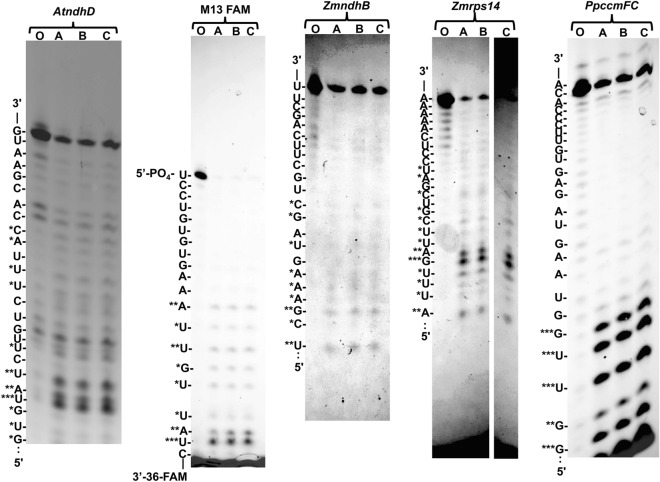
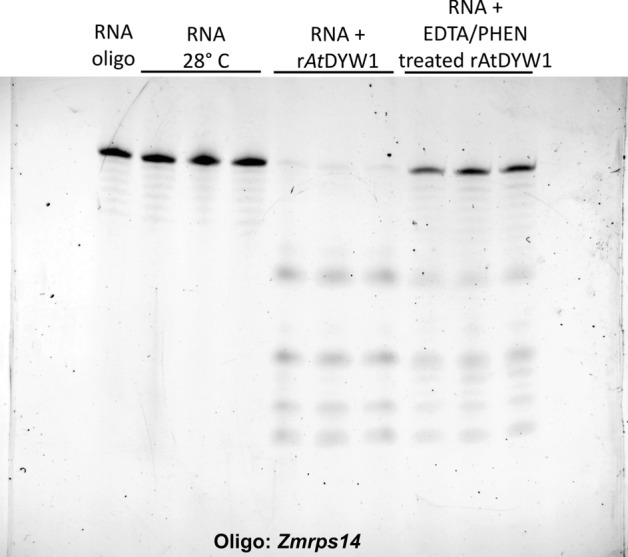

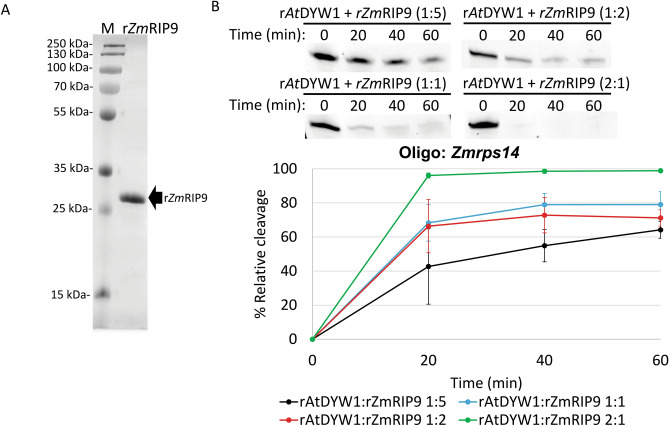
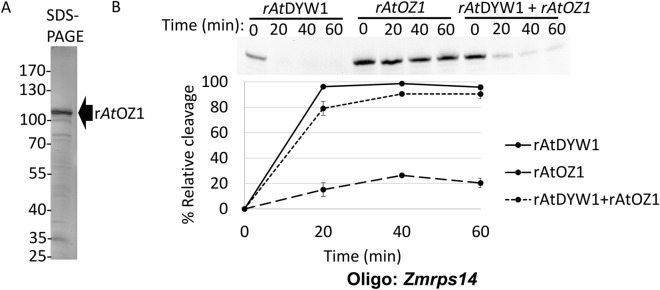
References
-
- Lurin C, Andrés C, Aubourg S, Bellaoui M, Bitton F, Bruyère C, Caboche M, Debast C, Gualberto J, Hoffmann B, et al. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell. 2004;16(8):2089–2103. doi: 10.1105/tpc.104.022236. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
