

Potent and uniform fetal hemoglobin induction via base editing
- PMID: 37400614
- PMCID: PMC10722557
- DOI: 10.1038/s41588-023-01434-7
Potent and uniform fetal hemoglobin induction via base editing
Abstract
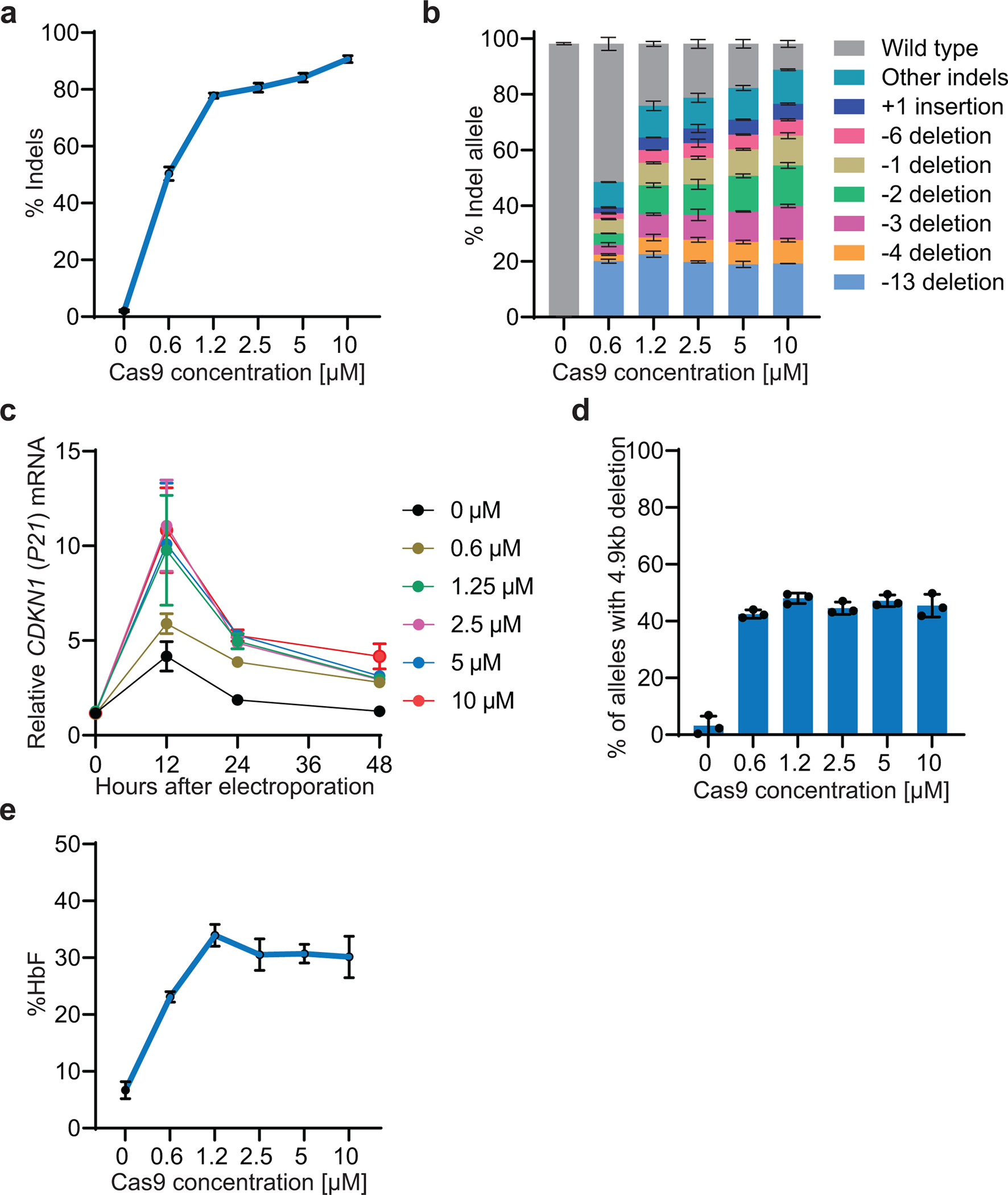
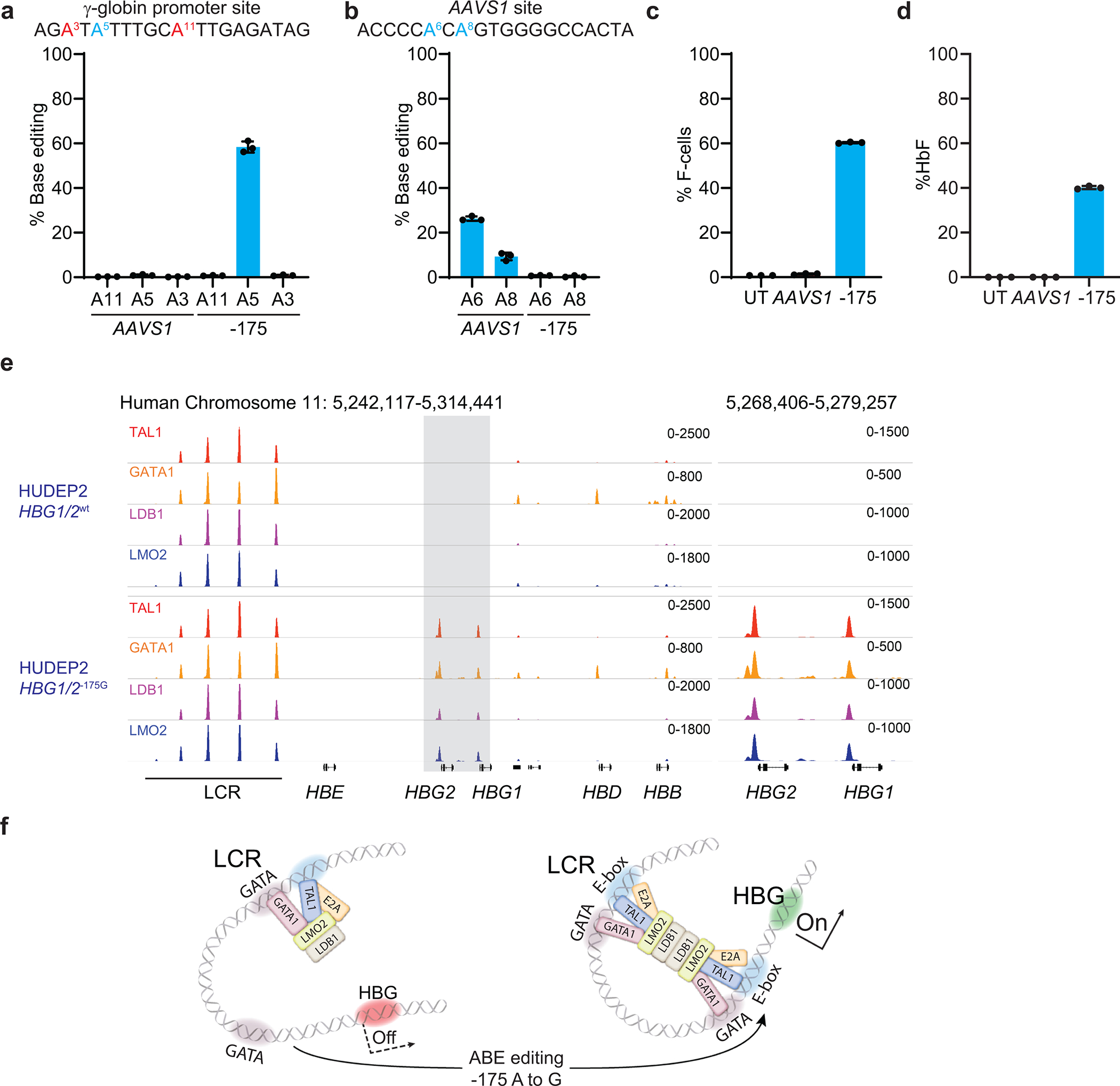
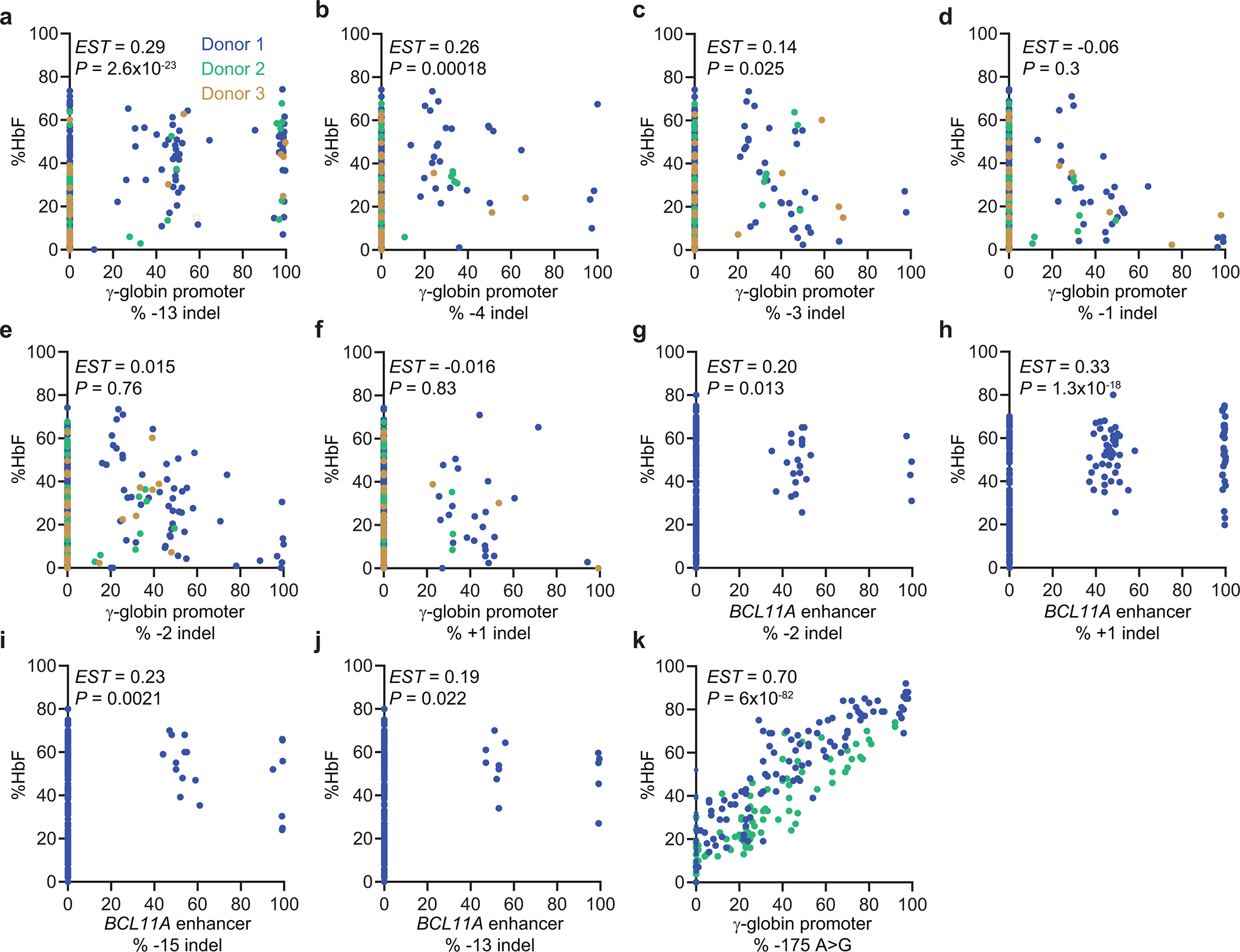
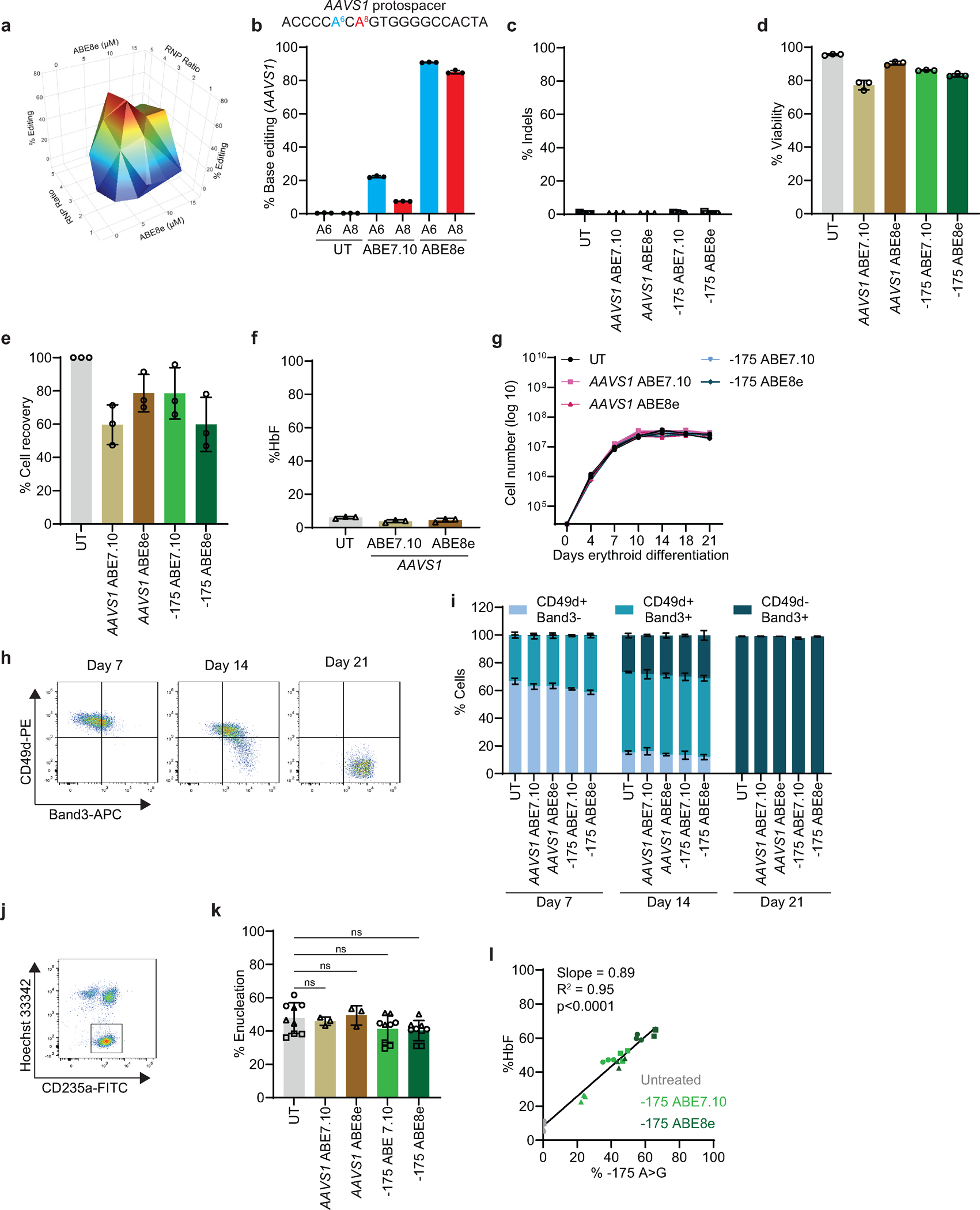
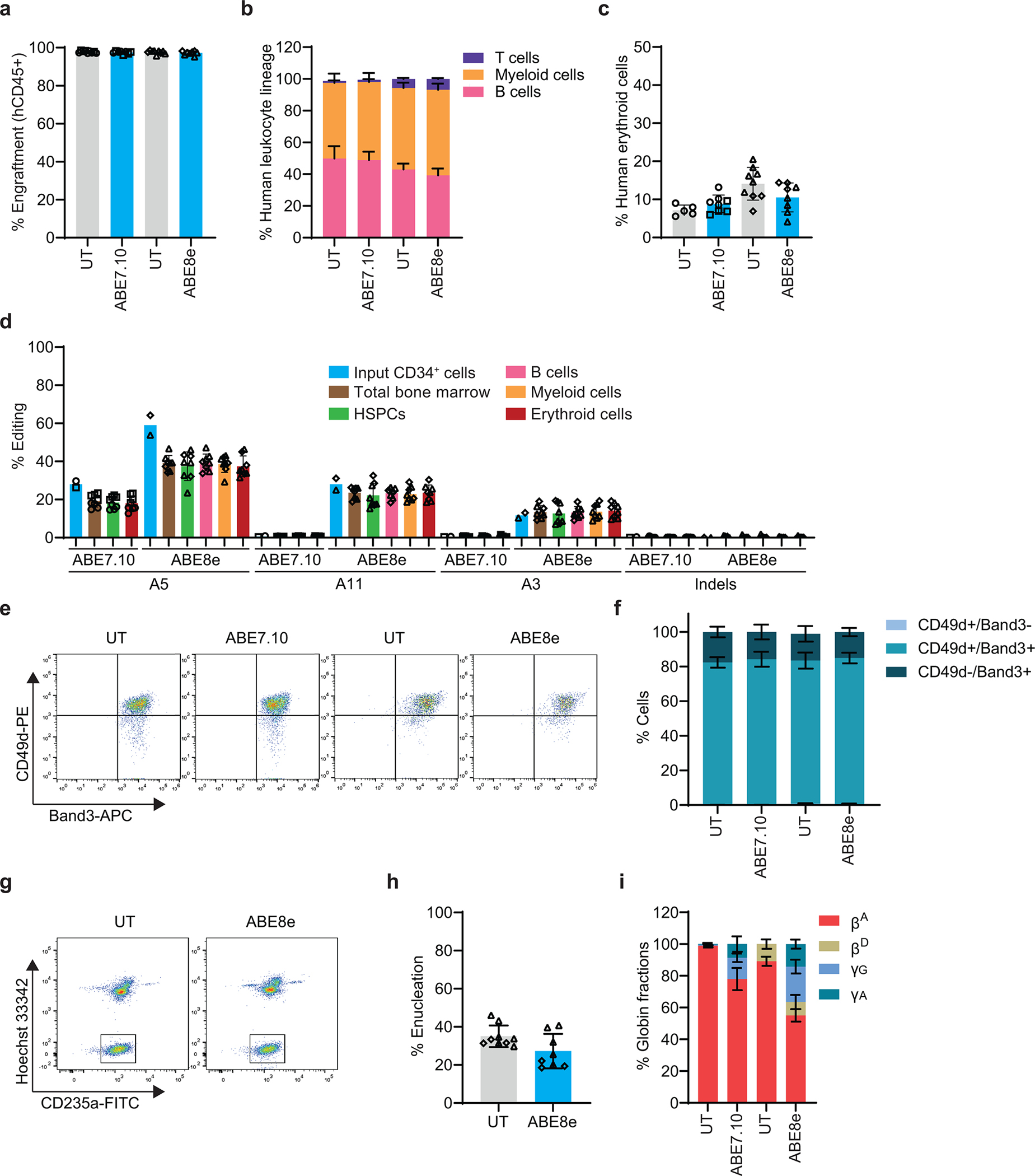
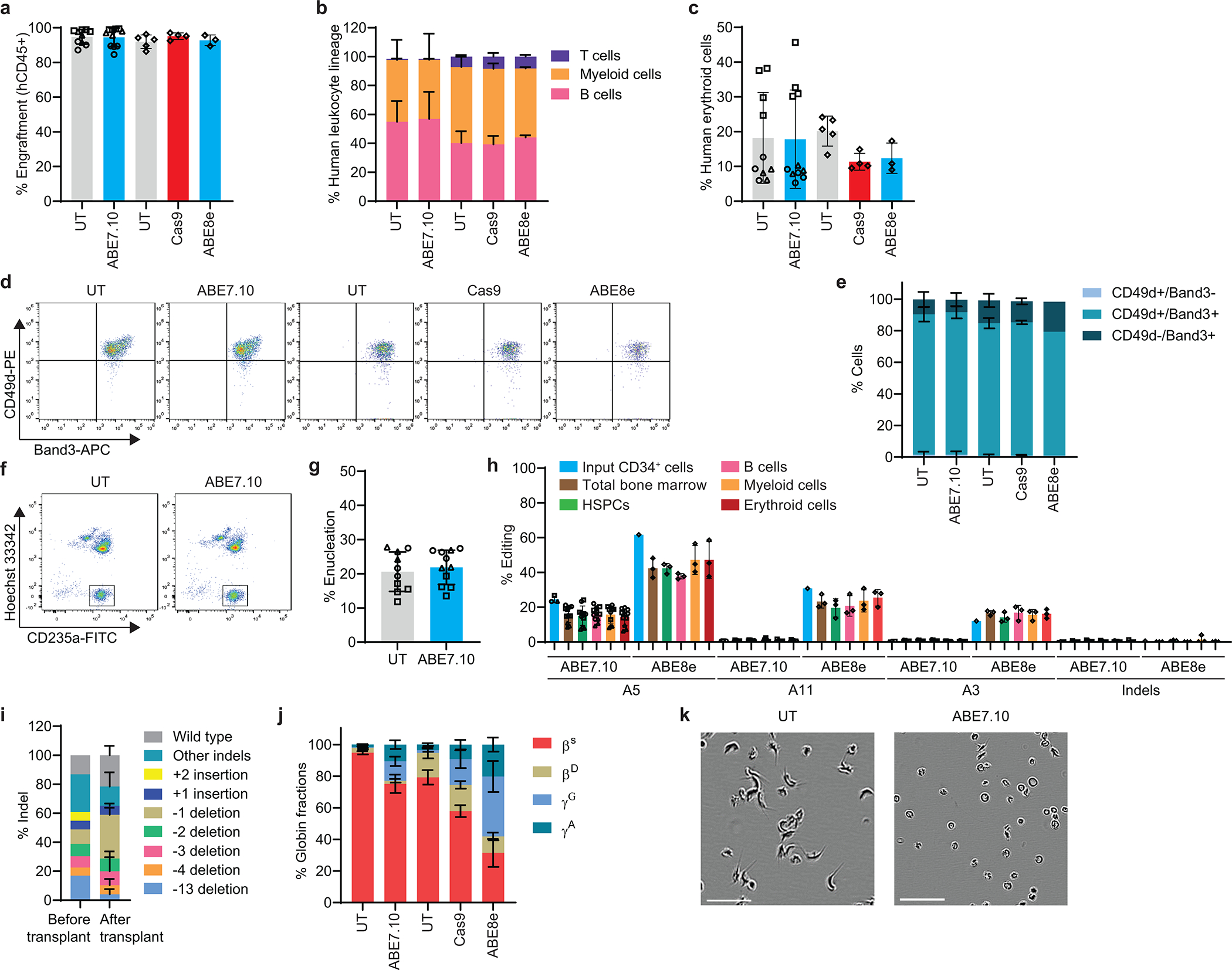
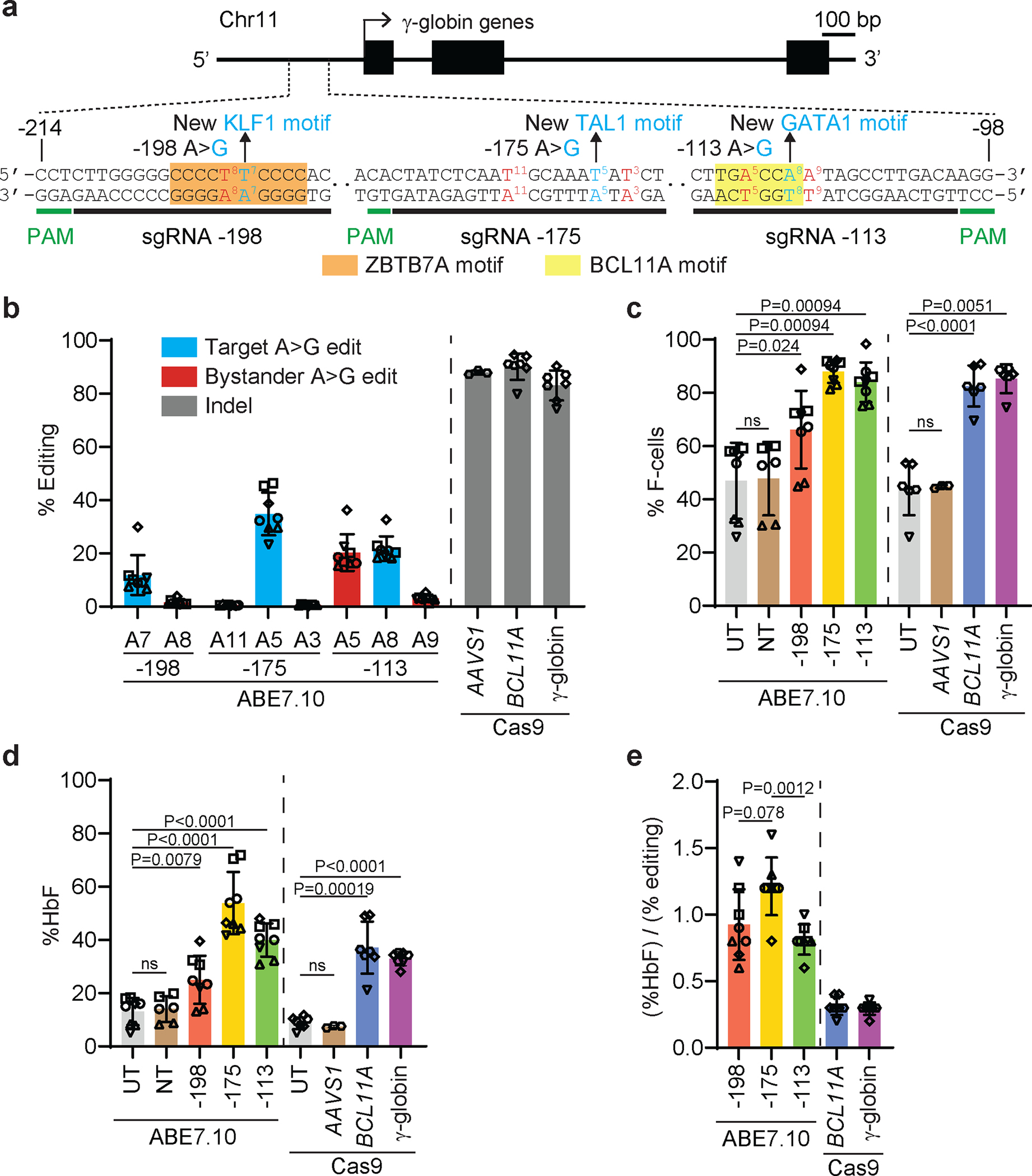
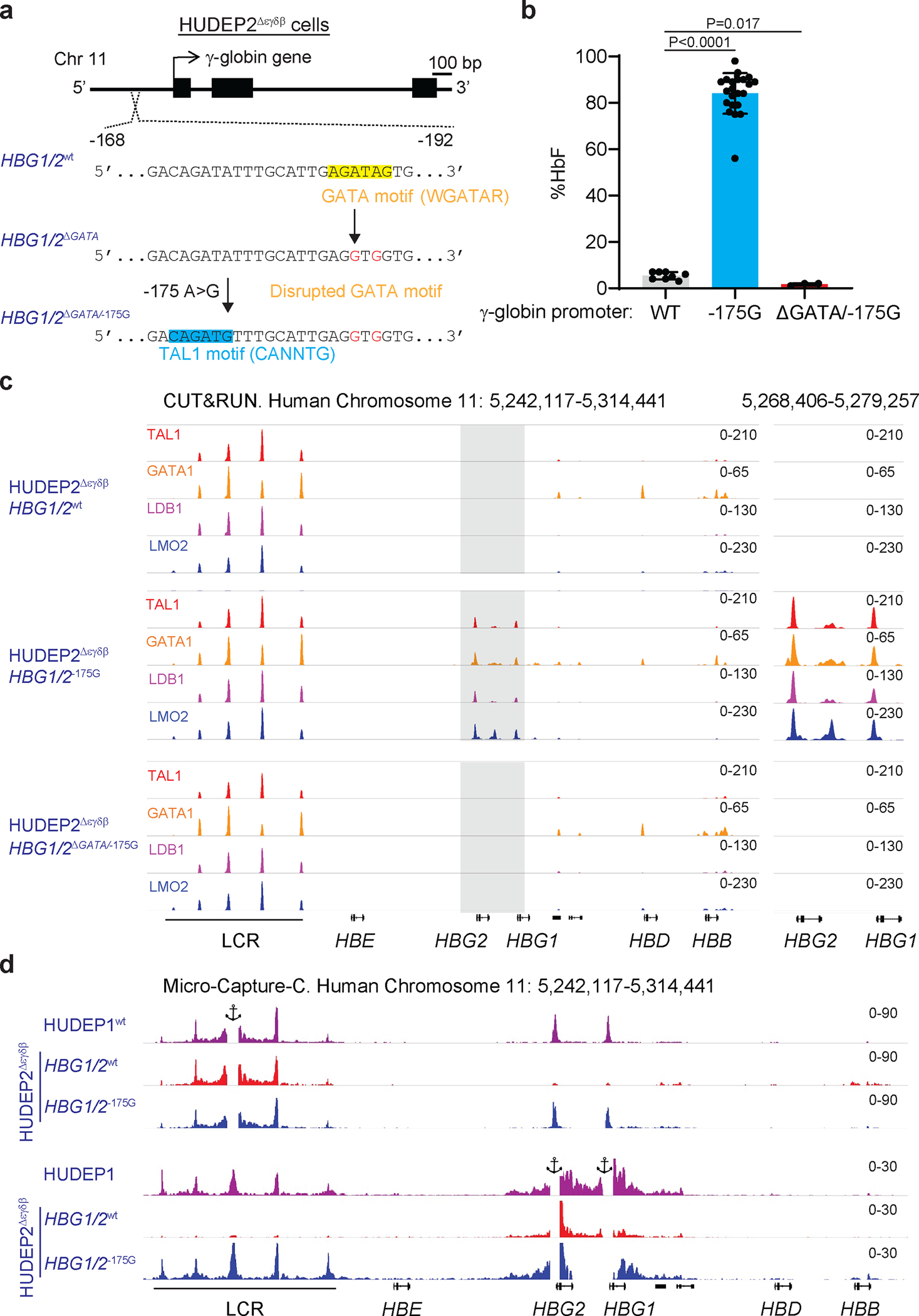
Inducing fetal hemoglobin (HbF) in red blood cells can alleviate β-thalassemia and sickle cell disease. We compared five strategies in CD34+ hematopoietic stem and progenitor cells, using either Cas9 nuclease or adenine base editors. The most potent modification was adenine base editor generation of γ-globin -175A>G. Homozygous -175A>G edited erythroid colonies expressed 81 ± 7% HbF versus 17 ± 11% in unedited controls, whereas HbF levels were lower and more variable for two Cas9 strategies targeting a BCL11A binding motif in the γ-globin promoter or a BCL11A erythroid enhancer. The -175A>G base edit also induced HbF more potently than a Cas9 approach in red blood cells generated after transplantation of CD34+ hematopoietic stem and progenitor cells into mice. Our data suggest a strategy for potent, uniform induction of HbF and provide insights into γ-globin gene regulation. More generally, we demonstrate that diverse indels generated by Cas9 can cause unexpected phenotypic variation that can be circumvented by base editing.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors have filed patent applications on genome-editing agents. D.R.L. is a consultant and equity owner of Beam Therapeutics, Prime Medicine, Pairwise Plants, and Chroma Medicine, companies that use genome editing or genome engineering. M.J.W. is a consultant for GSK plc, Cellarity Inc., Novartis, and Dyne Therapeutics. A.S. is a consultant for Spotlight Therapeutics, Medexus Inc., and Vertex Pharmaceuticals. A.S. has also received research funding from CRISPR Therapeutics and honoraria from Vindico Medical Education. A.S. is the St. Jude Children’s Research Hospital site principal investigator of clinical trials for genome editing of sickle cell disease sponsored by Vertex Pharmaceuticals/CRISPR Therapeutics (
Figures
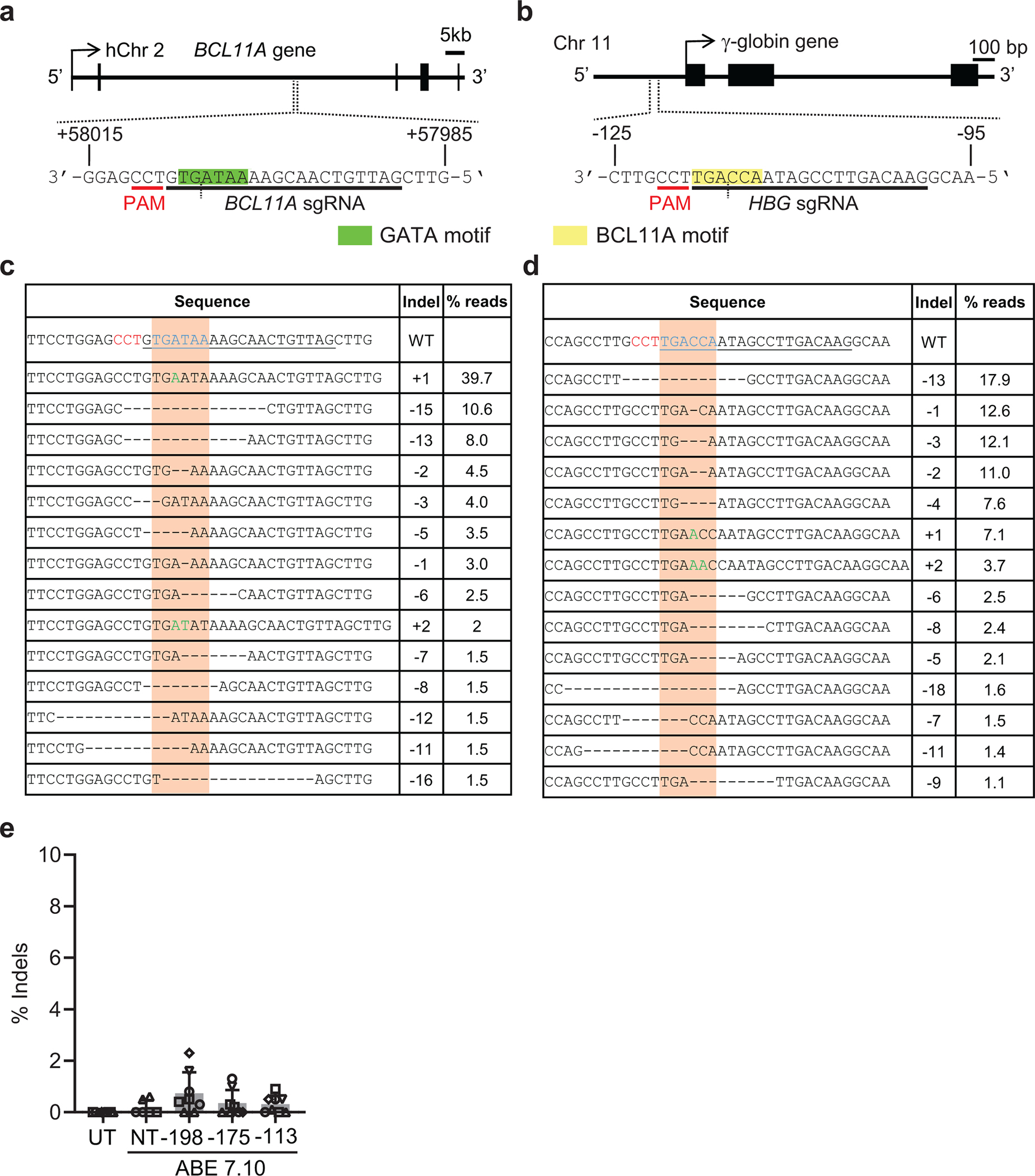
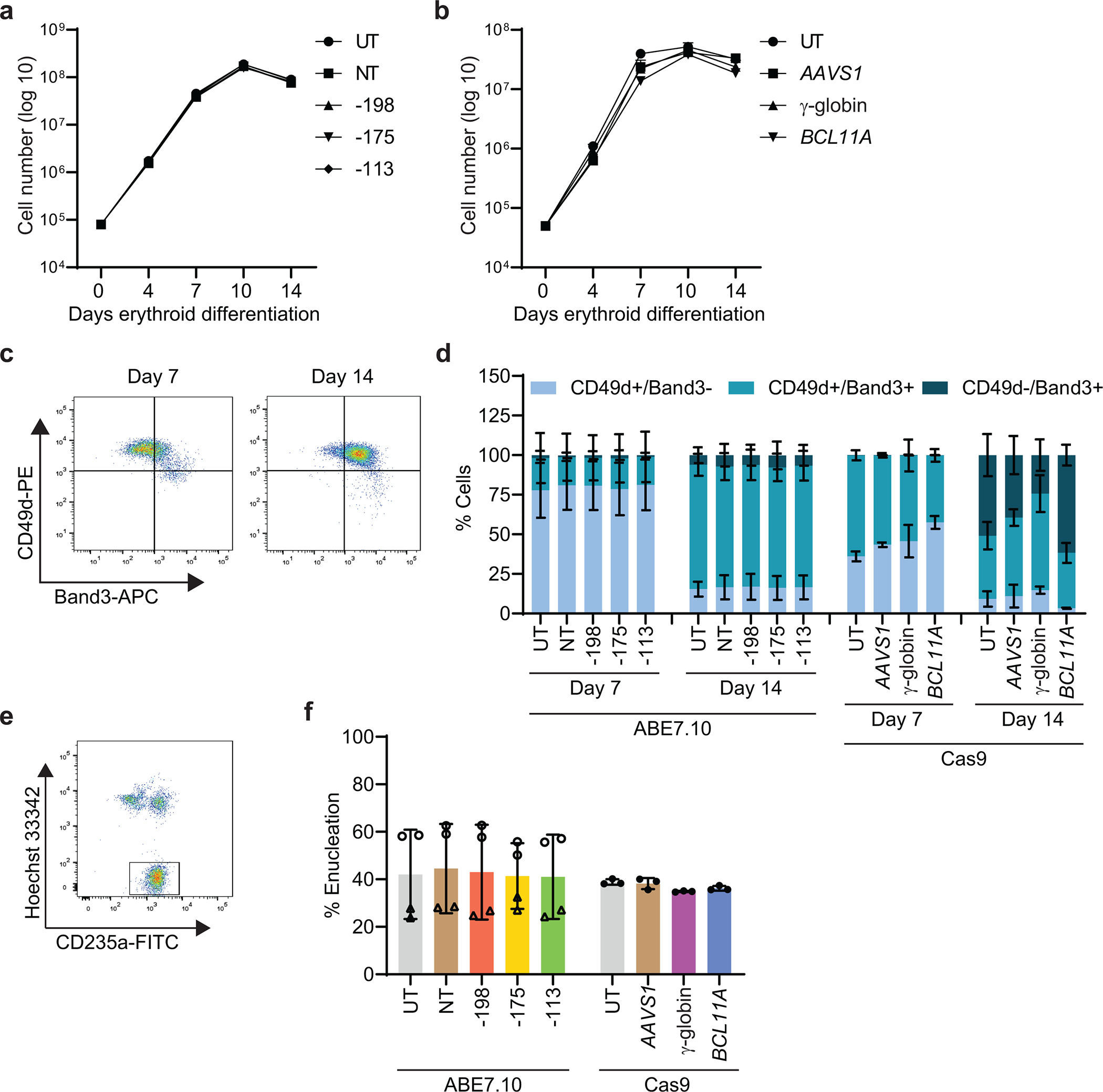
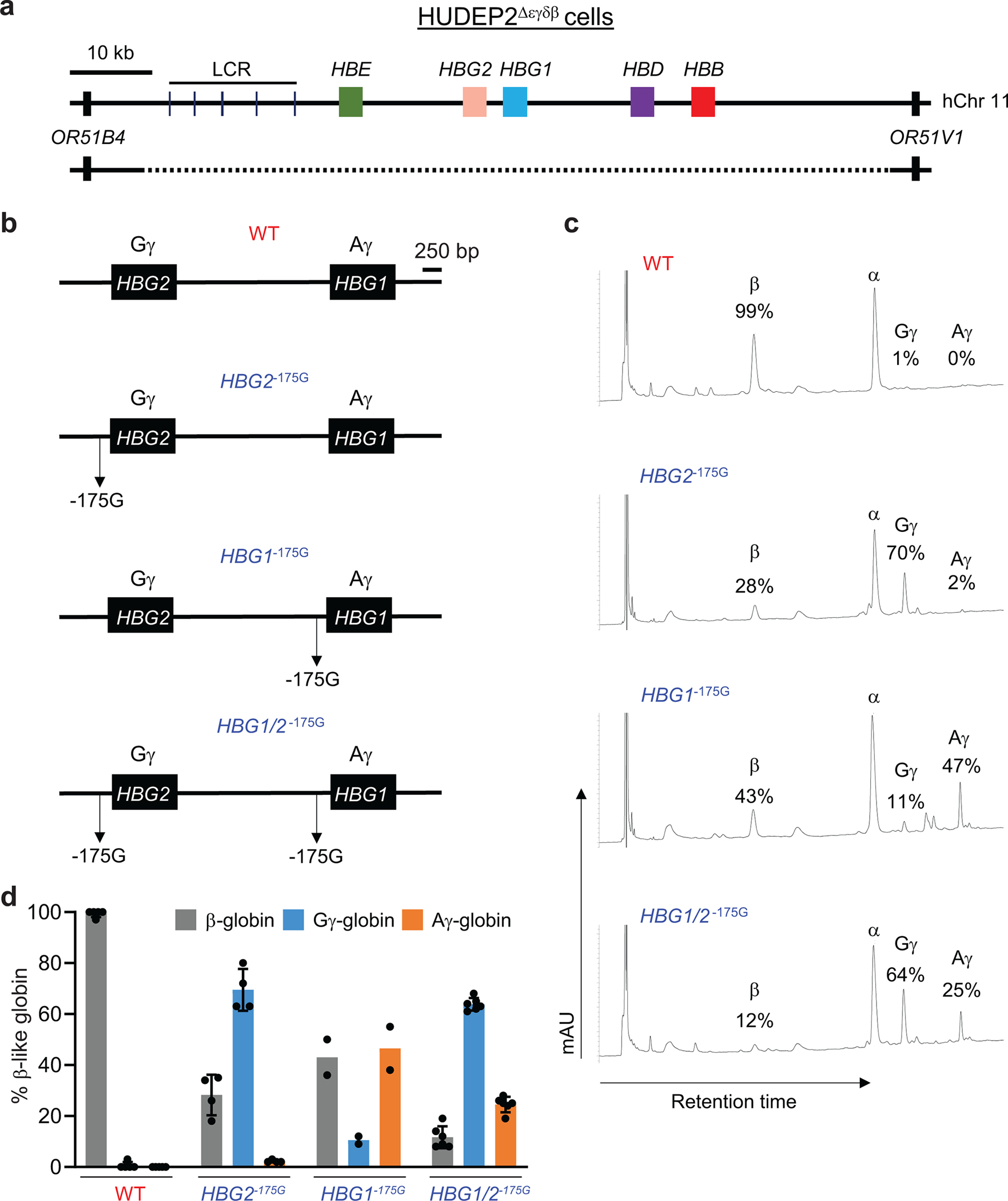
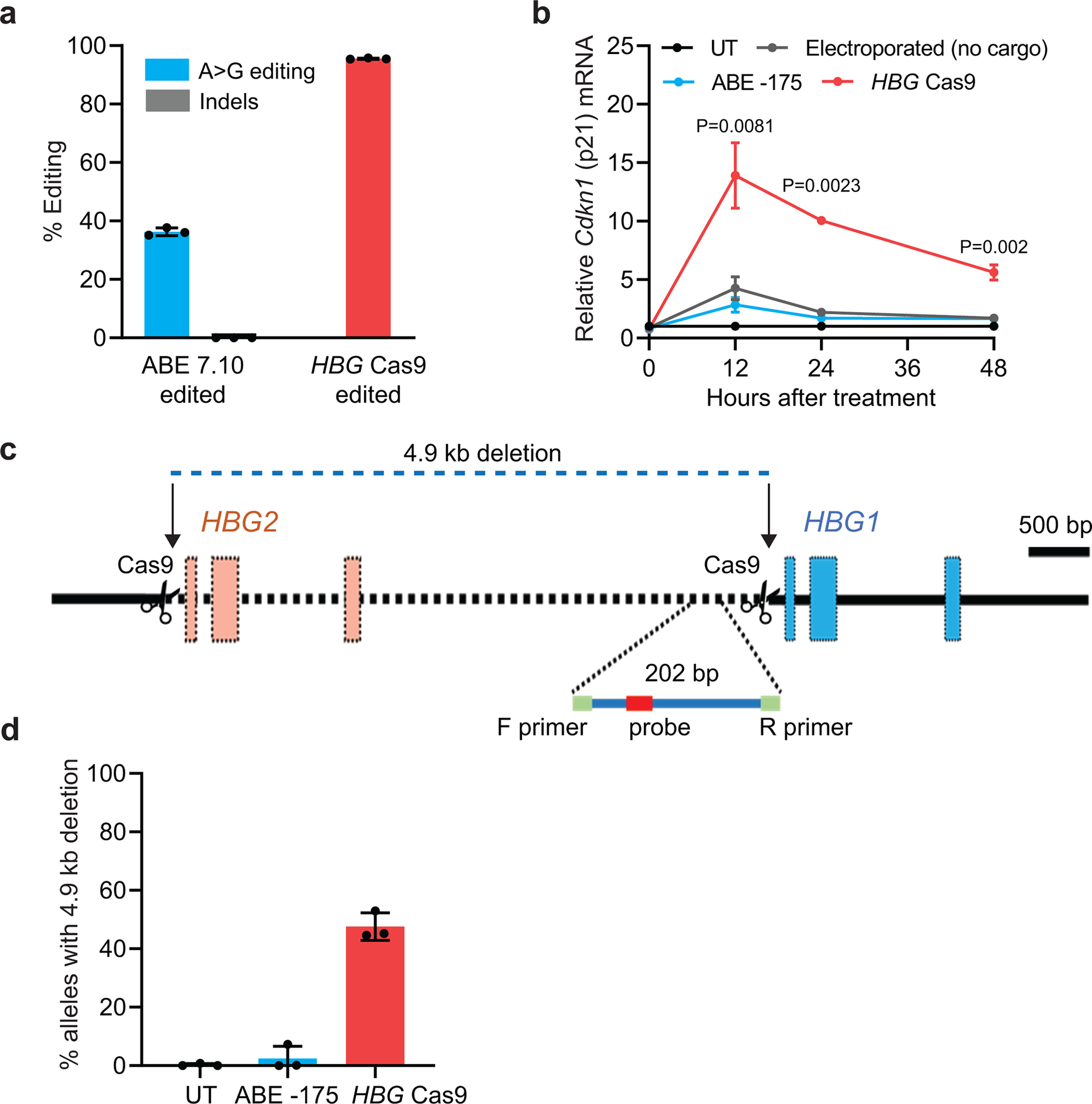
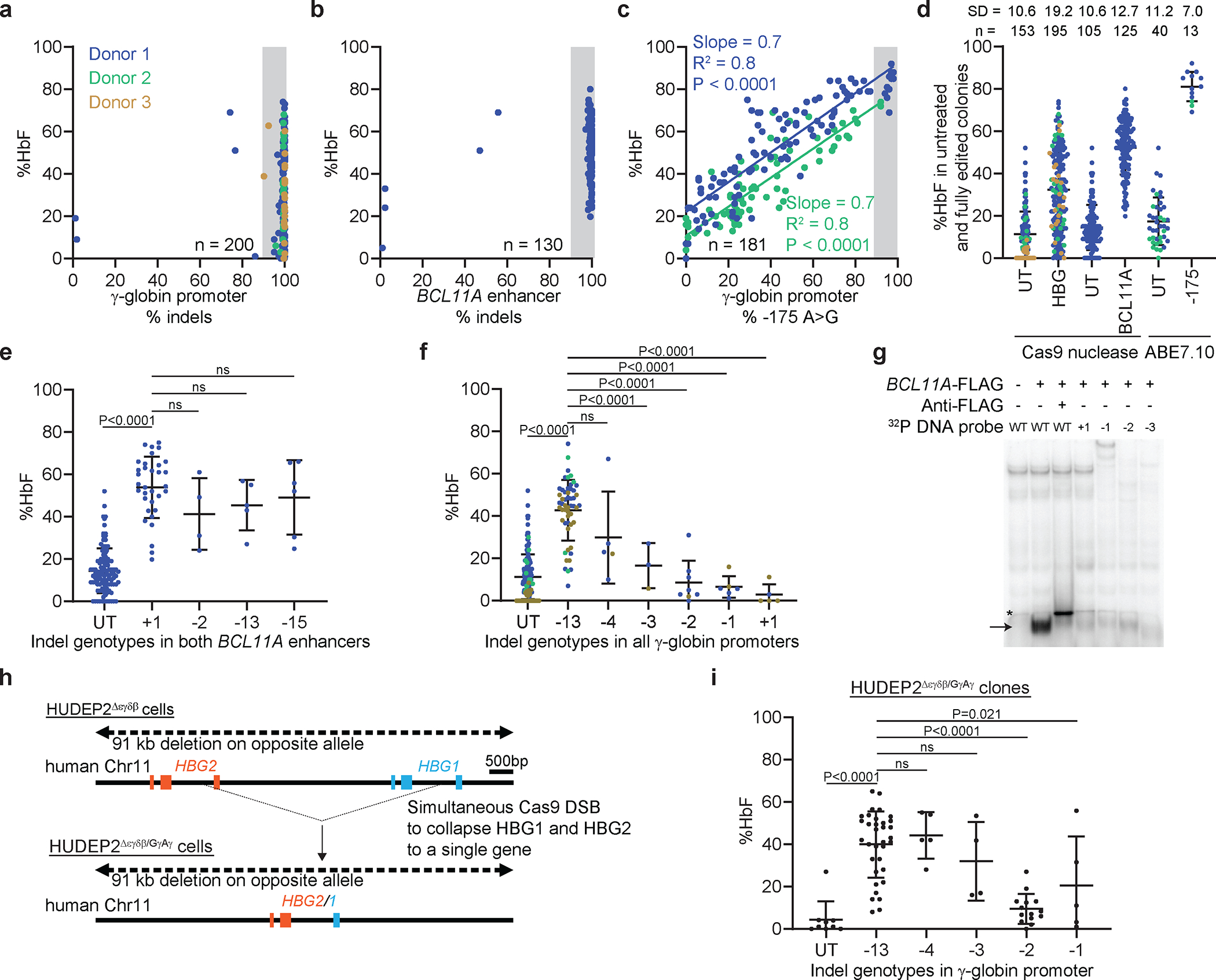
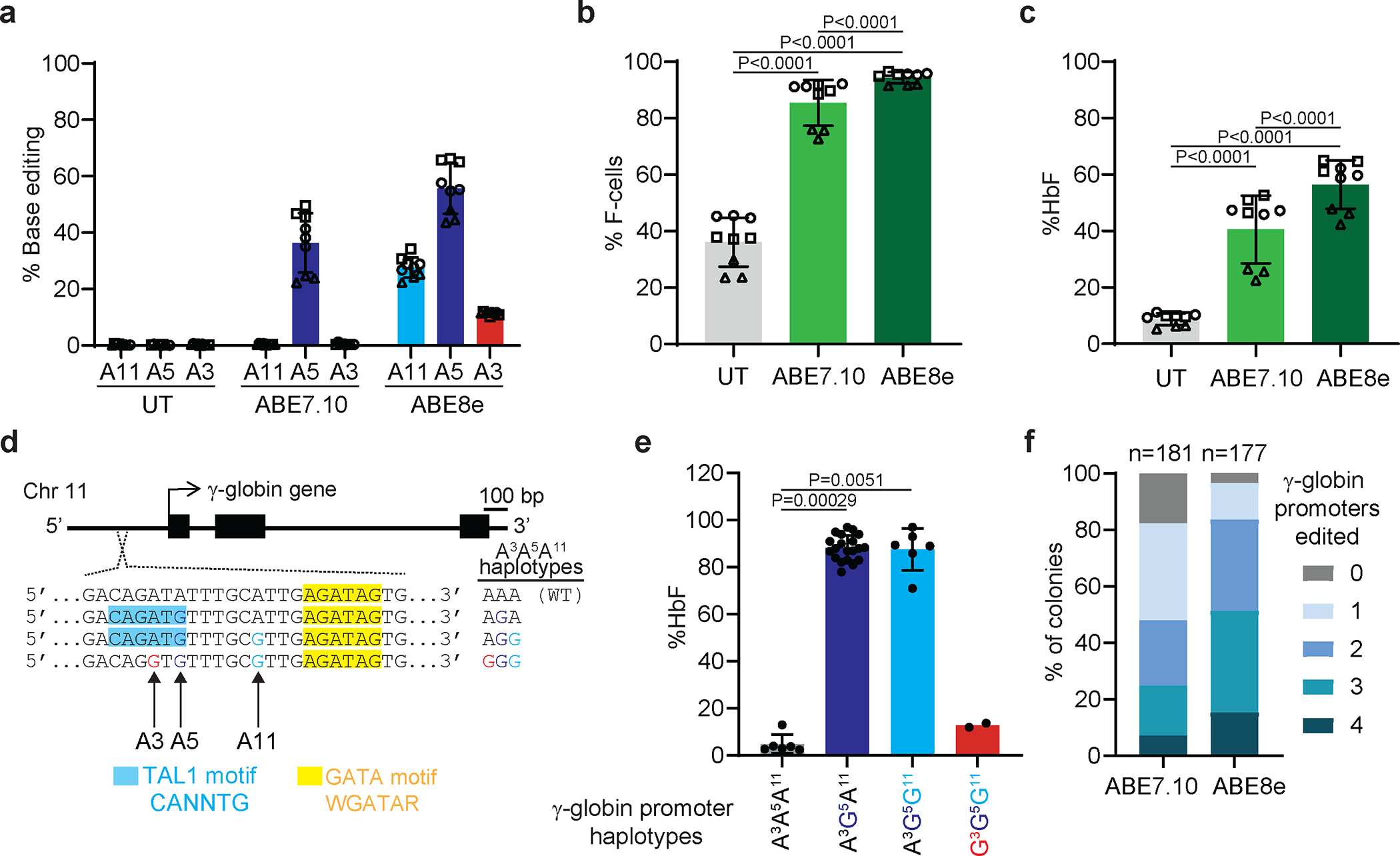
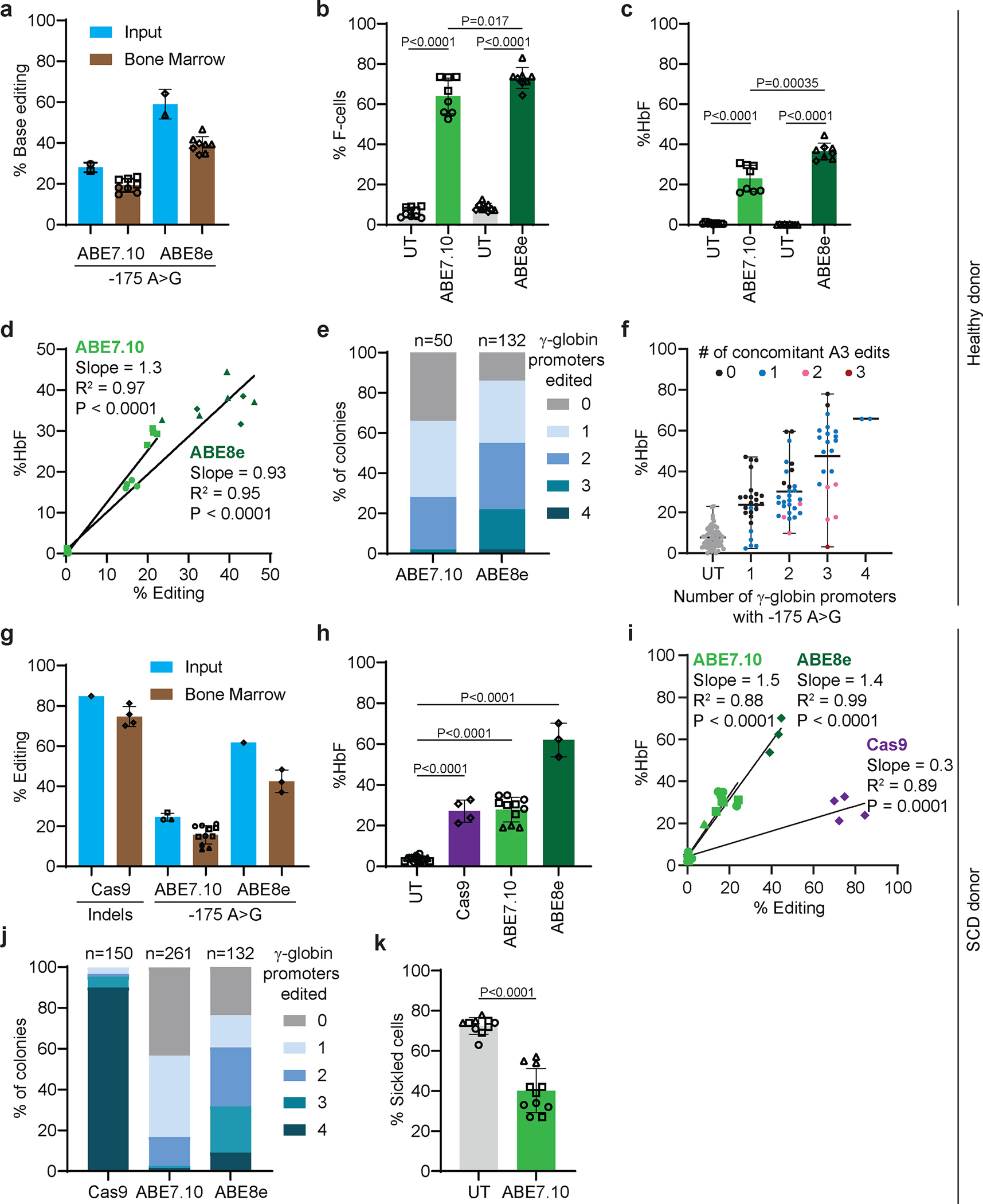
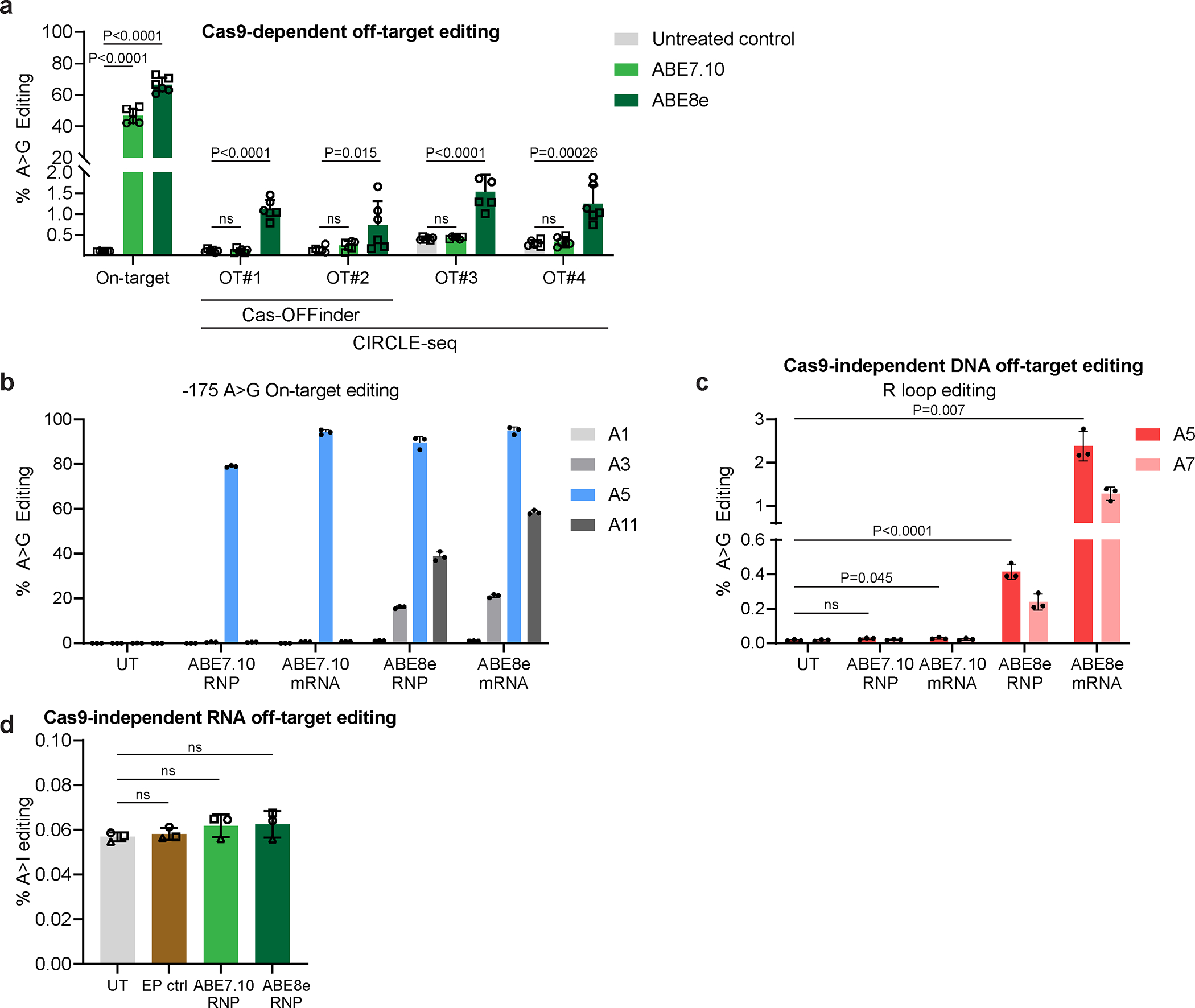
References
-
- Piel FB, Steinberg MH & Rees DC Sickle Cell Disease. N Engl J Med 376, 1561–1573 (2017). - PubMed
-
- Taher AT, Musallam KM & Cappellini MD beta-Thalassemias. N Engl J Med 384, 727–743 (2021). - PubMed
-
- Frangoul H et al. CRISPR-Cas9 Gene Editing for Sickle Cell Disease and β-Thalassemia. N. Engl. J. Med. 384, 252–260 (2021). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K99 HL163805/HL/NHLBI NIH HHS/United States
- R00 HL163805/HL/NHLBI NIH HHS/United States
- R35 GM118062/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P30 CA021765/CA/NCI NIH HHS/United States
- U01 AI142756/AI/NIAID NIH HHS/United States
- U01 AI157189/AI/NIAID NIH HHS/United States
- R01 EB022376/EB/NIBIB NIH HHS/United States
- R35 GM133614/GM/NIGMS NIH HHS/United States
- K01 DK132453/DK/NIDDK NIH HHS/United States
- R01 HL156647/HL/NHLBI NIH HHS/United States
- P01 HL053749/HL/NHLBI NIH HHS/United States
- R01 HL136135/HL/NHLBI NIH HHS/United States
- RM1 HG009490/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
