

Exploration of novel αβ-protein folds through de novo design
- PMID: 37400653
- PMCID: PMC10442233
- DOI: 10.1038/s41594-023-01029-0
Exploration of novel αβ-protein folds through de novo design
Abstract
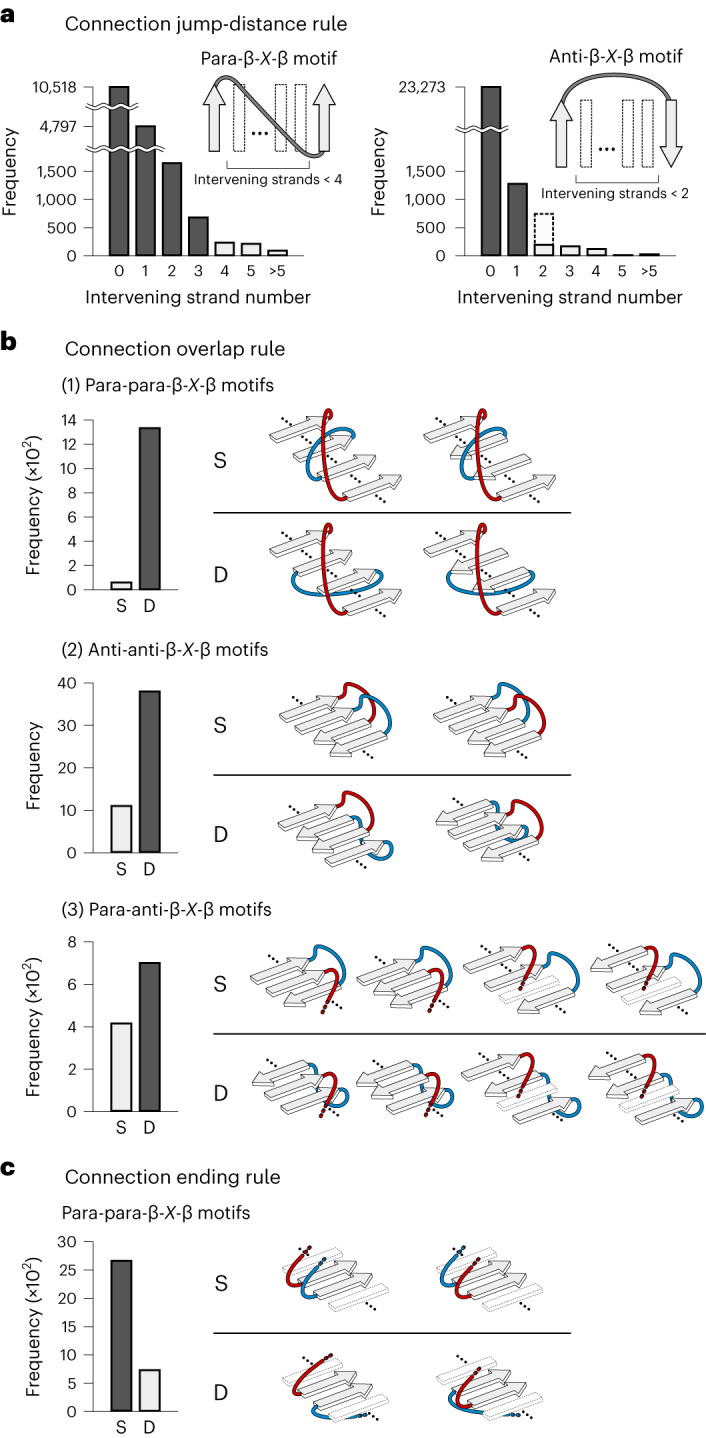
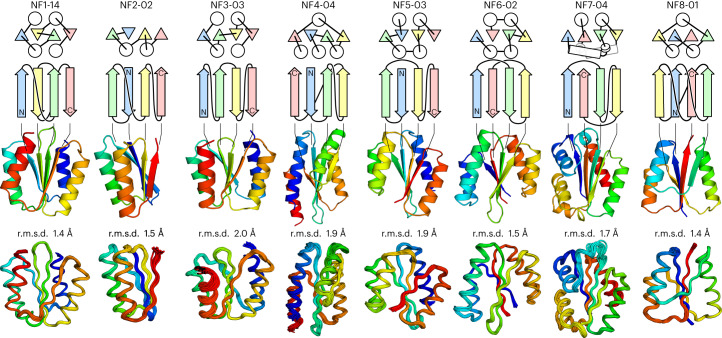
A fundamental question in protein evolution is whether nature has exhaustively sampled nearly all possible protein folds throughout evolution, or whether a large fraction of the possible folds remains unexplored. To address this question, we defined a set of rules for β-sheet topology to predict novel αβ-folds and carried out a systematic de novo protein design exploration of the novel αβ-folds predicted by the rules. The designs for all eight of the predicted novel αβ-folds with a four-stranded β-sheet, including a knot-forming one, folded into structures close to the design models. Further, the rules predicted more than 10,000 novel αβ-folds with five- to eight-stranded β-sheets; this number far exceeds the number of αβ-folds observed in nature so far. This result suggests that a vast number of αβ-folds are possible, but have not emerged or have become extinct due to evolutionary bias.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures












References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
