

Overexpression of human alpha-Synuclein leads to dysregulated microbiome/metabolites with ageing in a rat model of Parkinson disease
- PMID: 37403161
- PMCID: PMC10318726
- DOI: 10.1186/s13024-023-00628-1
Overexpression of human alpha-Synuclein leads to dysregulated microbiome/metabolites with ageing in a rat model of Parkinson disease
Abstract
Background: Braak's hypothesis states that sporadic Parkinson's disease (PD) follows a specific progression of pathology from the peripheral to the central nervous system, and this progression can be monitored by detecting the accumulation of alpha-Synuclein (α-Syn) protein. Consequently, there is growing interest in understanding how the gut (commensal) microbiome can regulate α-Syn accumulation, as this could potentially lead to PD.
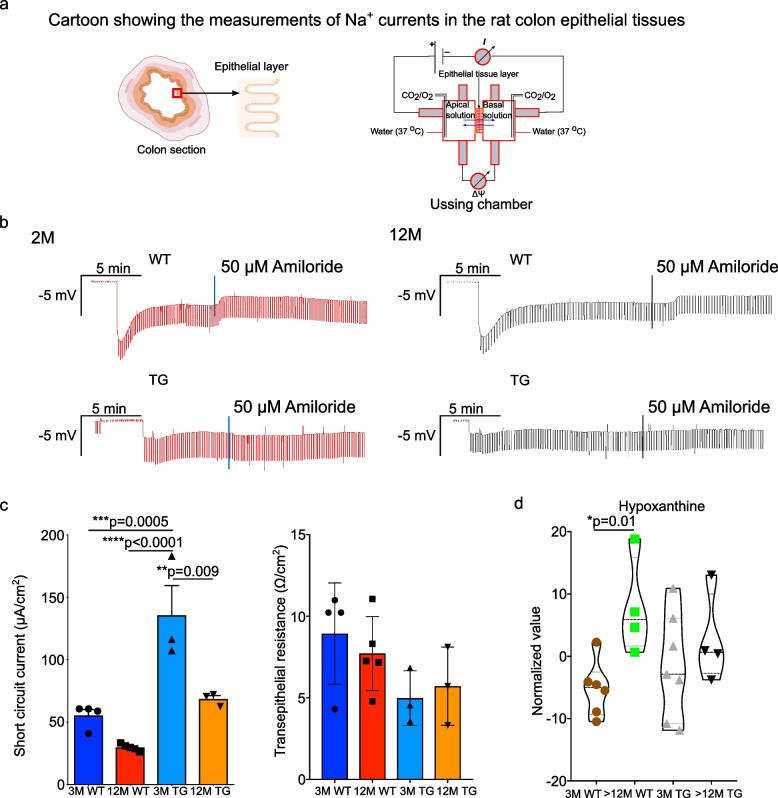
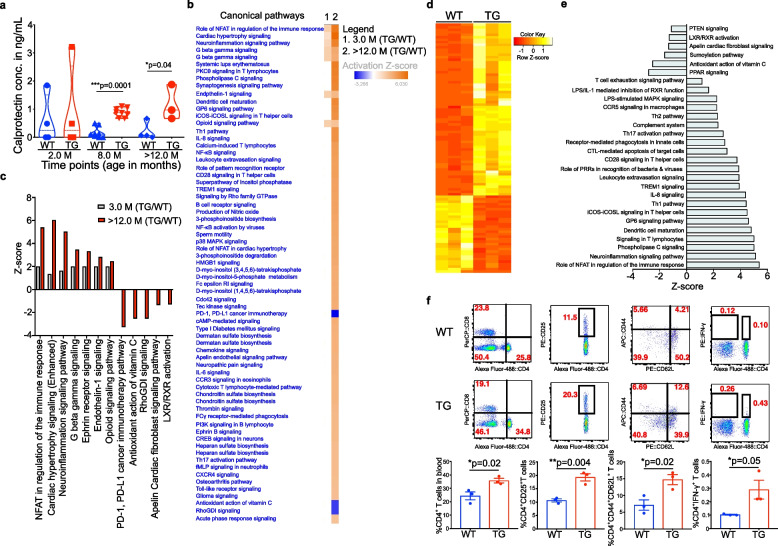
Methods: We used 16S rRNA and shotgun sequencing to characterise microbial diversity. 1H-NMR was employed to understand the metabolite production and intestinal inflammation estimated using ELISA and RNA-sequencing from feces and the intestinal epithelial layer respectively. The Na+ channel current and gut permeability were measured using an Ussing chamber. Immunohistochemistry and immunofluorescence imaging were applied to detect the α-Syn protein. LC-MS/MS was used for characterization of proteins from metabolite treated neuronal cells. Finally, Metascape and Ingenuity Pathway Analysis (IPA) bioinformatics tools were used for identification of dysregulated pathways.
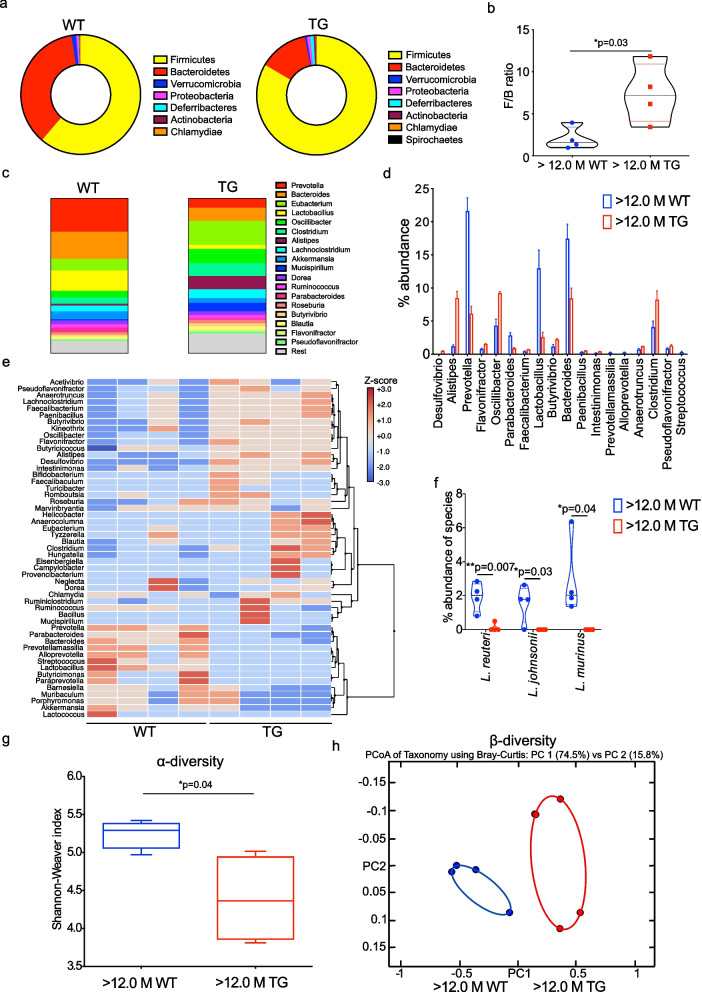
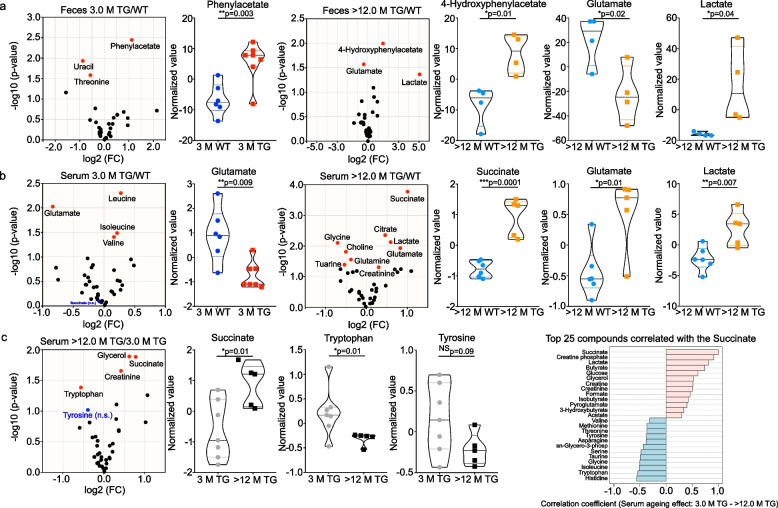
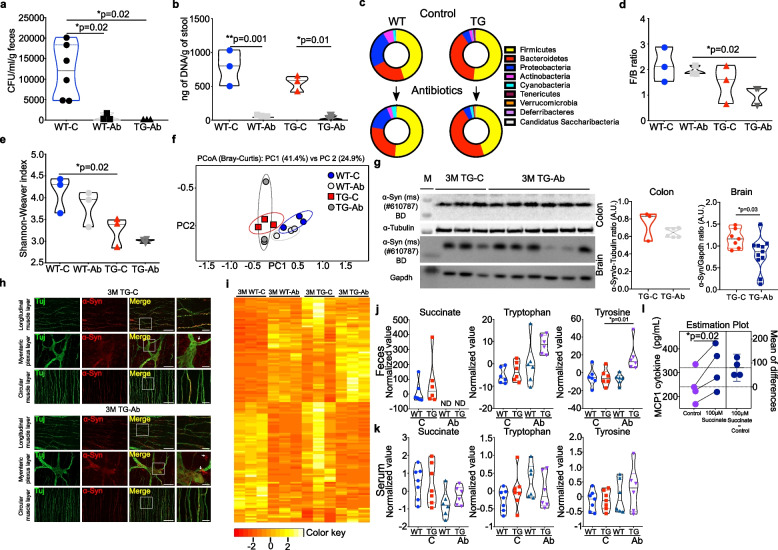
Results: We studied a transgenic (TG) rat model overexpressing the human SNCA gene and found that a progressive gut microbial composition alteration characterized by the reduction of Firmicutes to Bacteroidetes ratio could be detected in the young TG rats. Interestingly, this ratio then increased with ageing. The dynamics of Lactobacillus and Alistipes were monitored and reduced Lactobacillus and increased Alistipes abundance was discerned in ageing TG rats. Additionally, the SNCA gene overexpression resulted in gut α-Syn protein expression and increased with advanced age. Further, older TG animals had increased intestinal inflammation, decreased Na+ current and a robust alteration in metabolite production characterized by the increase of succinate levels in feces and serum. Manipulation of the gut bacteria by short-term antibiotic cocktail treatment revealed a complete loss of short-chain fatty acids and a reduction in succinate levels. Although antibiotic cocktail treatment did not change α-Syn expression in the enteric nervous system of the colon, however, reduced α-Syn expression was detected in the olfactory bulbs (forebrain) of the TG rats.
Conclusion: Our data emphasize that the gut microbiome dysbiosis synchronous with ageing leads to a specific alteration of gut metabolites and can be modulated by antibiotics which may affect PD pathology.
Keywords: Antibiotics; Gut microbiome; Intestinal inflammation; PD; α-Synuclein.
© 2023. The Author(s).
Conflict of interest statement
All the authors declared that they have no competing interest.
Figures


References
-
- Antunes L, Frasquilho S, Ostaszewski M, Weber J, Longhino L, Antony P, Baumuratov A, Buttini M, Shannon KM, Balling R, et al. Similar alpha-Synuclein staining in the colon mucosa in patients with Parkinson's disease and controls. Mov Disord. 2016;31(10):1567–1570. doi: 10.1002/mds.26702. - DOI - PubMed
-
- Zhong CB, Chen QQ, Haikal C, Li W, Svanbergsson A, Diepenbroek M, Li JY. Age-dependent alpha-synuclein accumulation and phosphorylation in the enteric nervous system in a transgenic mouse model of Parkinson's Disease. Neurosci Bull. 2017;33(5):483–492. doi: 10.1007/s12264-017-0179-1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
