

The fission yeast methyl phosphate capping enzyme Bmc1 guides 2'-O-methylation of the U6 snRNA
- PMID: 37403782
- PMCID: PMC10484740
- DOI: 10.1093/nar/gkad563
The fission yeast methyl phosphate capping enzyme Bmc1 guides 2'-O-methylation of the U6 snRNA
Abstract
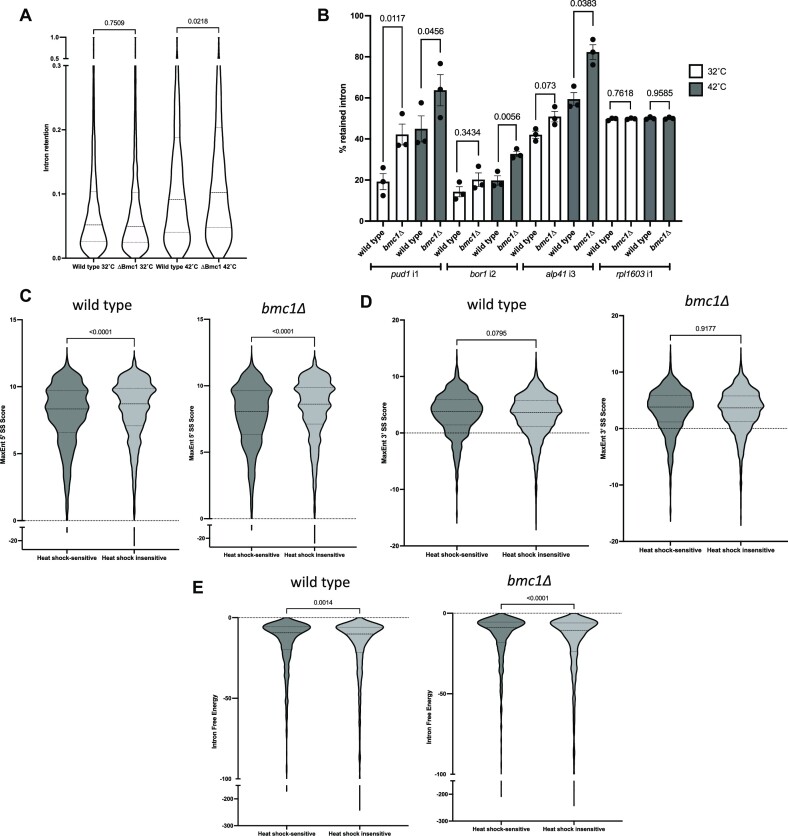
Splicing requires the tight coordination of dynamic spliceosomal RNAs and proteins. U6 is the only spliceosomal RNA transcribed by RNA Polymerase III and undergoes an extensive maturation process. In humans and fission yeast, this includes addition of a 5' γ-monomethyl phosphate cap by members of the Bin3/MePCE family as well as snoRNA guided 2'-O-methylation. Previously, we have shown that the Bin3/MePCE homolog Bmc1 is recruited to the S. pombe telomerase holoenzyme by the LARP7 family protein Pof8, where it acts in a catalytic-independent manner to protect the telomerase RNA and facilitate holoenzyme assembly. Here, we show that Bmc1 and Pof8 are required for the formation of a distinct U6 snRNP that promotes 2'-O-methylation of U6, and identify a non-canonical snoRNA that guides this methylation. We also show that the 5' γ-monomethyl phosphate capping activity of Bmc1 is not required for its role in promoting snoRNA guided 2'-O-methylation, and that this role relies on different regions of Pof8 from those required for Pof8 function in telomerase. Our results are consistent with a novel role for Bmc1/MePCE family members in stimulating 2'-O-methylation and a more general role for Bmc1 and Pof8 in guiding noncoding RNP assembly beyond the telomerase RNP.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






Similar articles
-
The methyl phosphate capping enzyme Bmc1/Bin3 is a stable component of the fission yeast telomerase holoenzyme.Nat Commun. 2022 Mar 11;13(1):1277. doi: 10.1038/s41467-022-28985-3. Nat Commun. 2022. PMID: 35277511 Free PMC article.
-
A putative cap binding protein and the methyl phosphate capping enzyme Bin3/MePCE function in telomerase biogenesis.Nat Commun. 2022 Feb 25;13(1):1067. doi: 10.1038/s41467-022-28545-9. Nat Commun. 2022. PMID: 35217638 Free PMC article.
-
The Schizosaccharomyces pombe mgU6-47 gene is required for 2'-O-methylation of U6 snRNA at A41.Nucleic Acids Res. 2002 Feb 15;30(4):894-902. doi: 10.1093/nar/30.4.894. Nucleic Acids Res. 2002. PMID: 11842100 Free PMC article.
-
RNA modification in Cajal bodies.RNA Biol. 2017 Jun 3;14(6):693-700. doi: 10.1080/15476286.2016.1249091. Epub 2016 Oct 24. RNA Biol. 2017. PMID: 27775477 Free PMC article. Review.
-
U6 RNA biogenesis and disease association.Wiley Interdiscip Rev RNA. 2013 Sep-Oct;4(5):581-92. doi: 10.1002/wrna.1181. Epub 2013 Jun 14. Wiley Interdiscip Rev RNA. 2013. PMID: 23776162 Review.
Cited by
-
Key RNA-binding domains in the La protein establish tRNA modification levels in Trypanosoma brucei.Nucleic Acids Res. 2025 Jul 8;53(13):gkaf594. doi: 10.1093/nar/gkaf594. Nucleic Acids Res. 2025. PMID: 40637228 Free PMC article.
-
Circuit logic: interdependent RNA modifications shape mRNA and noncoding RNA structure and function.RNA. 2025 Apr 16;31(5):613-622. doi: 10.1261/rna.080421.125. RNA. 2025. PMID: 40044218 Free PMC article. Review.
-
Unlocking the life code: a review of SnoRNA functional diversity and disease relevance.Cell Commun Signal. 2025 Jun 4;23(1):266. doi: 10.1186/s12964-025-02274-0. Cell Commun Signal. 2025. PMID: 40468441 Free PMC article. Review.
References
-
- Wilkinson M.E., Charenton C., Nagai K.. RNA splicing by the spliceosome. Annu. Rev. Biochem. 2020; 89:359–388. - PubMed
-
- Eysmont K., Matylla-Kuli Nska K., Jaskulska A., Magnus M., Konarska Correspondence M.M., Matylla-Kulińska K., Jaskulska A., Magnus M., Konarska M.M. Rearrangements within the U6 snRNA core during the transition between the two catalytic steps of splicing. Mol. Cell. 2019; 75:538–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
