

Peptidyl-prolyl cis/trans isomerase Pin1 interacts with hepatitis B virus core particle, but not with HBc protein, to promote HBV replication
- PMID: 37404723
- PMCID: PMC10315659
- DOI: 10.3389/fcimb.2023.1195063
Peptidyl-prolyl cis/trans isomerase Pin1 interacts with hepatitis B virus core particle, but not with HBc protein, to promote HBV replication
Abstract
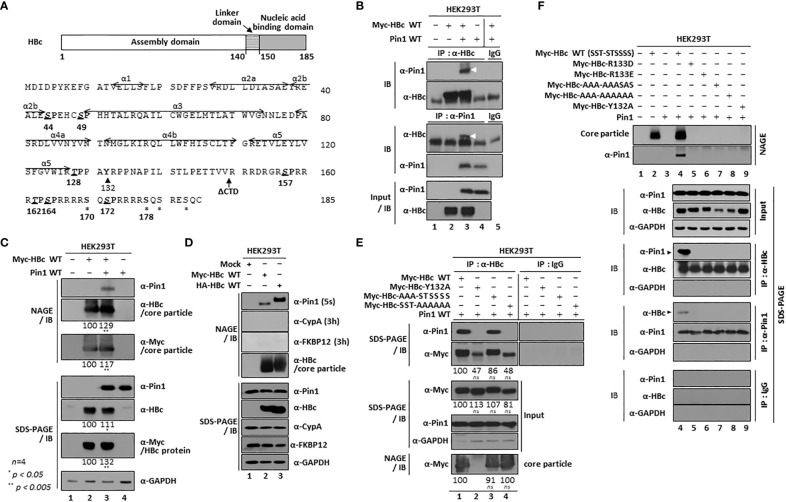
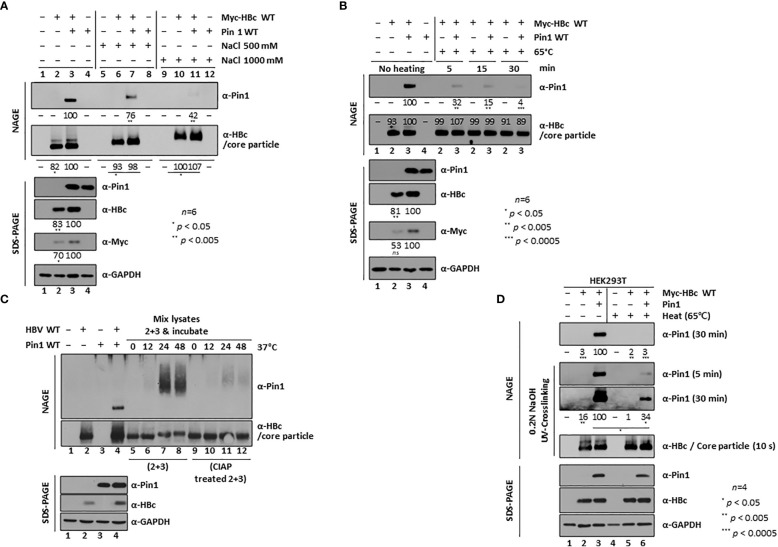
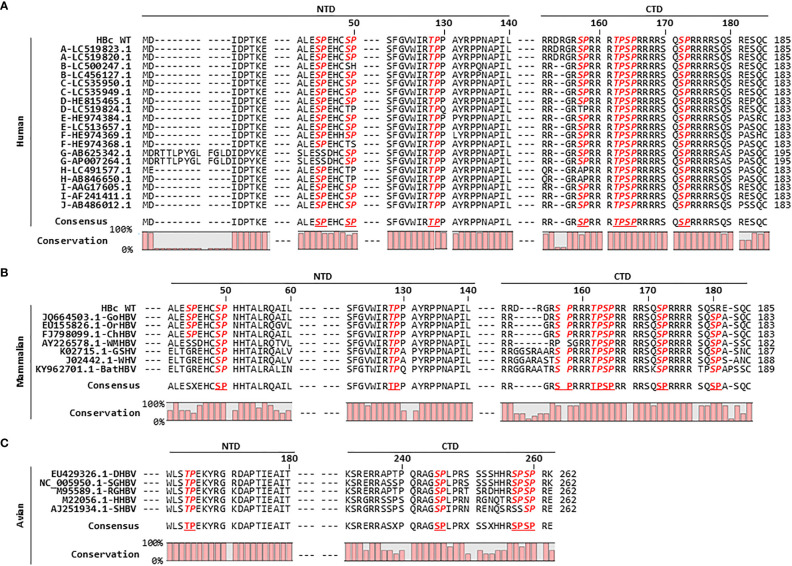
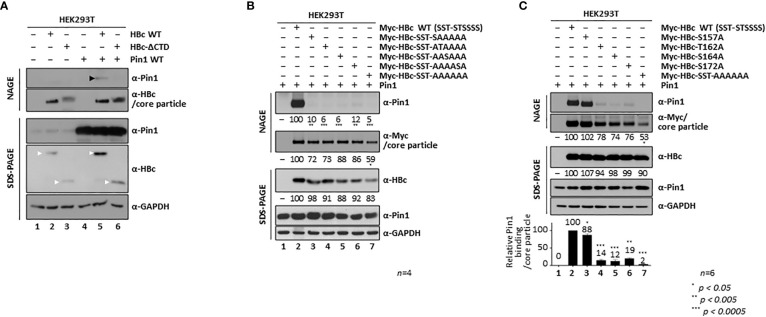
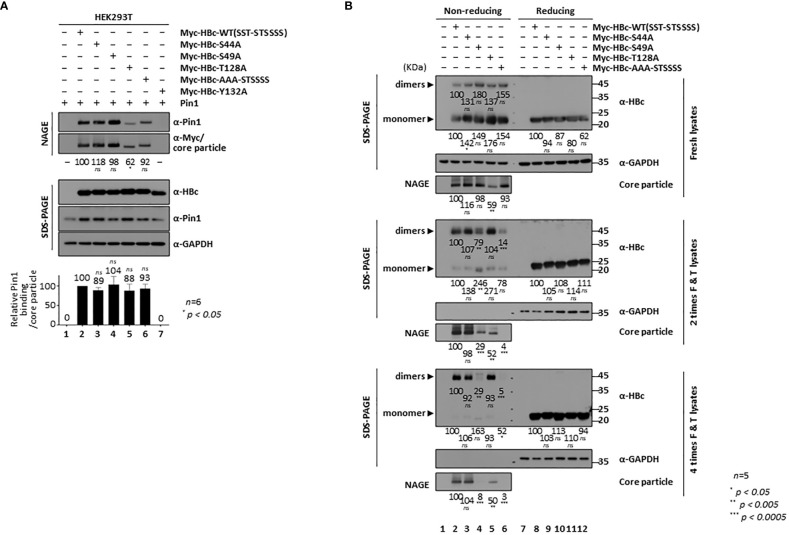
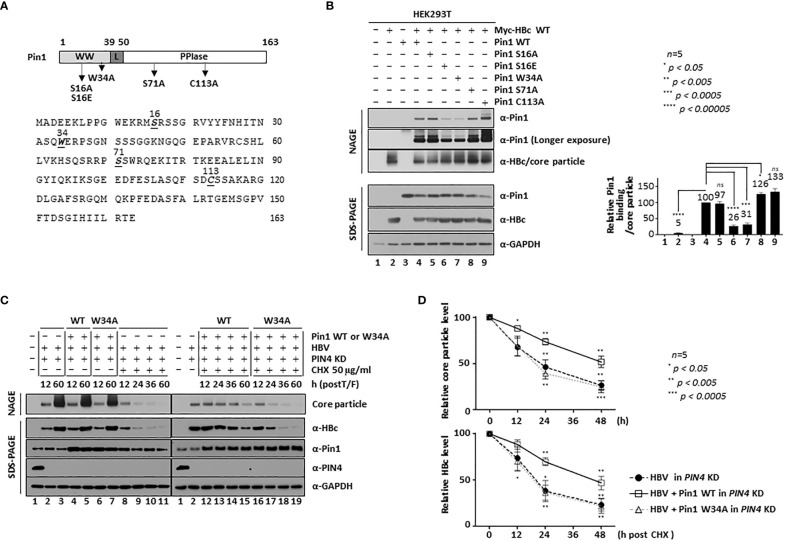
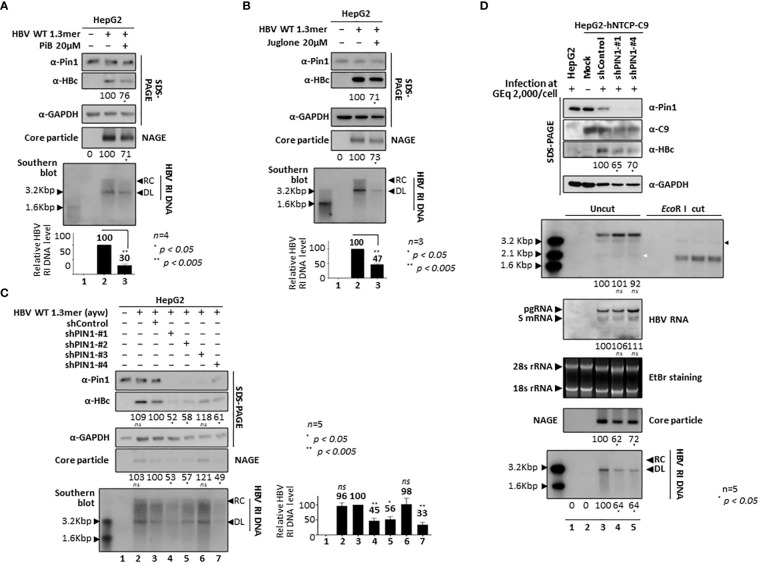
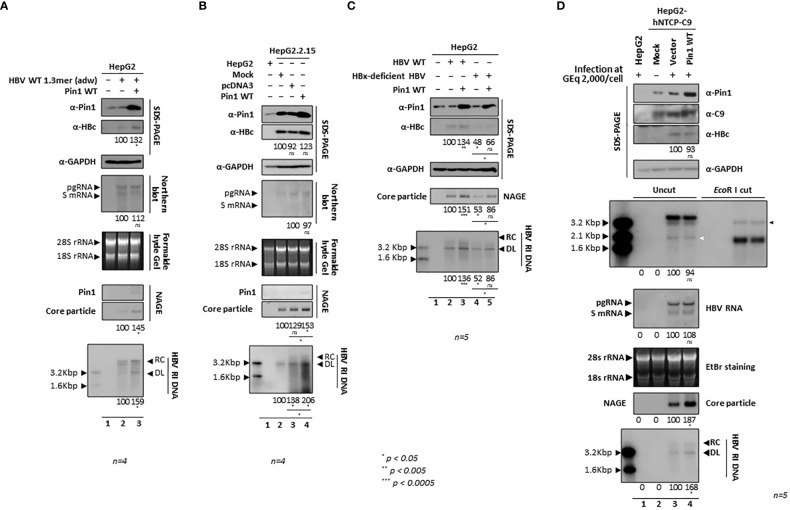
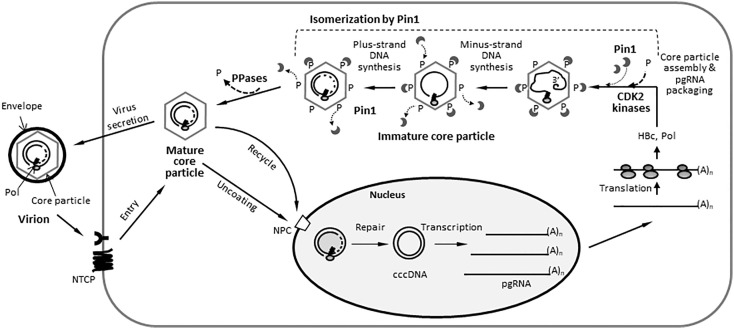
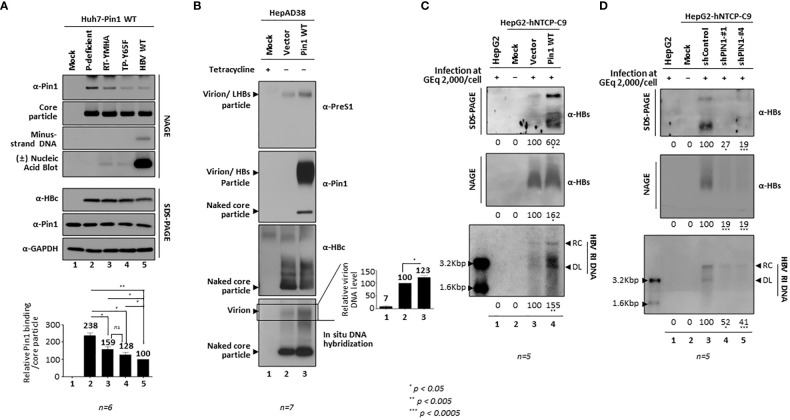
Here, we demonstrate that the peptidyl-prolyl cis/trans isomerase Pin1 interacts noncovalently with the hepatitis B virus (HBV) core particle through phosphorylated serine/threonine-proline (pS/TP) motifs in the carboxyl-terminal domain (CTD) but not with particle-defective, dimer-positive mutants of HBc. This suggests that neither dimers nor monomers of HBc are Pin1-binding partners. The 162TP, 164SP, and 172SP motifs within the HBc CTD are important for the Pin1/core particle interaction. Although Pin1 dissociated from core particle upon heat treatment, it was detected as an opened-up core particle, demonstrating that Pin1 binds both to the outside and the inside of the core particle. Although the amino-terminal domain S/TP motifs of HBc are not involved in the interaction, 49SP contributes to core particle stability, and 128TP might be involved in core particle assembly, as shown by the decreased core particle level of S49A mutant through repeated freeze and thaw and low-level assembly of the T128A mutant, respectively. Overexpression of Pin1 increased core particle stability through their interactions, HBV DNA synthesis, and virion secretion without concomitant increases in HBV RNA levels, indicating that Pin1 may be involved in core particle assembly and maturation, thereby promoting the later stages of the HBV life cycle. By contrast, parvulin inhibitors and PIN1 knockdown reduced HBV replication. Since more Pin1 proteins bound to immature core particles than to mature core particles, the interaction appears to depend on the stage of virus replication. Taken together, the data suggest that physical association between Pin1 and phosphorylated core particles may induce structural alterations through isomerization by Pin1, induce dephosphorylation by unidentified host phosphatases, and promote completion of virus life cycle.
Keywords: HBV replication; PPIase Pin1; Pin1-core particle interaction; core particle; hepatitis B virus.
Copyright © 2023 Kwon, Kim, Song, Sajjad, Ha, Jung, Park, Shin and Kim.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous