

All-Covalent Nuclease-Resistant and Hydrogel-Tethered DNA Hairpin Probes Map pN Cell Traction Forces
- PMID: 37409737
- PMCID: PMC10360067
- DOI: 10.1021/acsami.3c04826
All-Covalent Nuclease-Resistant and Hydrogel-Tethered DNA Hairpin Probes Map pN Cell Traction Forces
Abstract
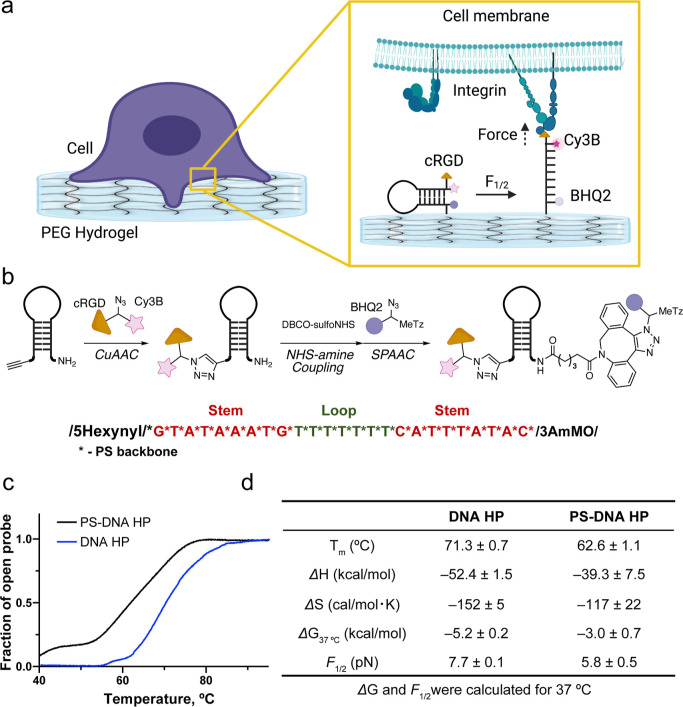
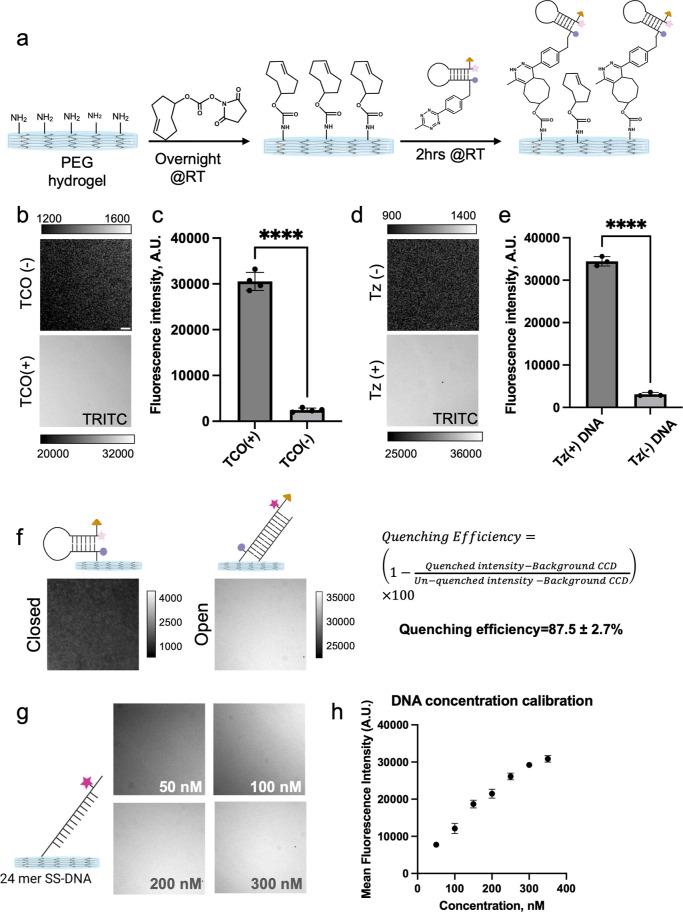
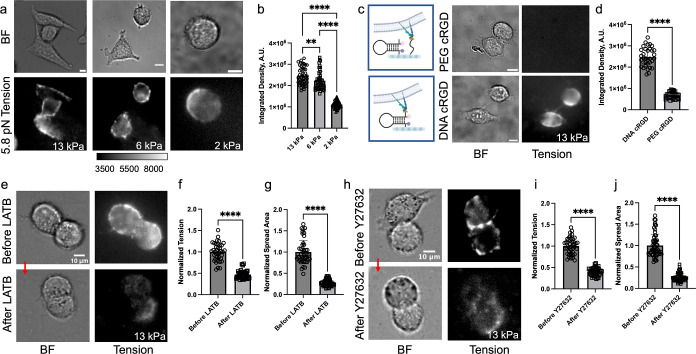
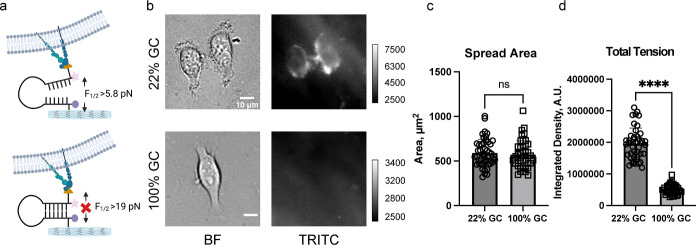
Cells sense and respond to the physical properties of their environment through receptor-mediated signaling, a process known as mechanotransduction, which can modulate critical cellular functions such as proliferation, differentiation, and survival. At the molecular level, cell adhesion receptors, such as integrins, transmit piconewton (pN)-scale forces to the extracellular matrix, and the magnitude of the force plays a critical role in cell signaling. The most sensitive approach to measuring integrin forces involves DNA hairpin-based sensors, which are used to quantify and map forces in living cells. Despite the broad use of DNA hairpin sensors to study a variety of mechanotransduction processes, these sensors are typically anchored to rigid glass slides, which are orders of magnitude stiffer than the extracellular matrix and hence modulate native biological responses. Here, we have developed nuclease-resistant DNA hairpin probes that are all covalently tethered to PEG hydrogels to image cell traction forces on physiologically relevant substrate stiffness. Using HeLa cells as a model cell line, we show that the molecular forces transmitted by integrins are highly sensitive to the bulk modulus of the substrate, and cells cultured on the 6 and 13 kPa gels produced a greater number of hairpin unfolding events compared to the 2 kPa substrates. Tension signals are spatially colocalized with pY118-paxillin, confirming focal adhesion-mediated probe opening. Additionally, we found that integrin forces are greater than 5.8 pN but less than 19 pN on 13 kPa gels. This work provides a general strategy to integrate molecular tension probes into hydrogels, which can better mimic in vivo mechanotransduction.
Keywords: DNA hairpin sensors; DNA tension probes; hydrogel; integrin mechanosensing; mechanobiology; mechanotransduction.
Conflict of interest statement
The authors declare no competing financial interest.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
