

Direct interaction of small non-coding RNAs CjNC140 and CjNC110 optimizes expression of key pathogenic phenotypes of Campylobacter jejuni
- PMID: 37409826
- PMCID: PMC10470494
- DOI: 10.1128/mbio.00833-23
Direct interaction of small non-coding RNAs CjNC140 and CjNC110 optimizes expression of key pathogenic phenotypes of Campylobacter jejuni
Abstract
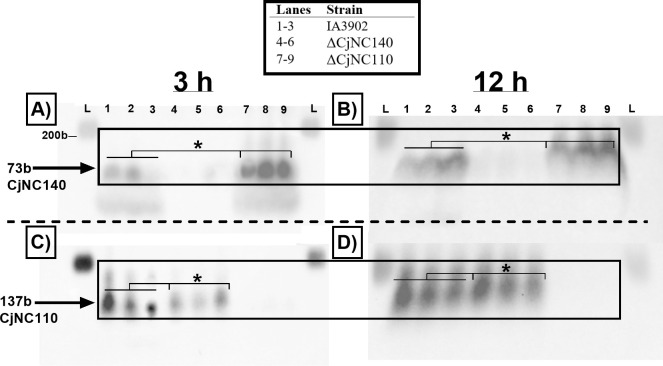
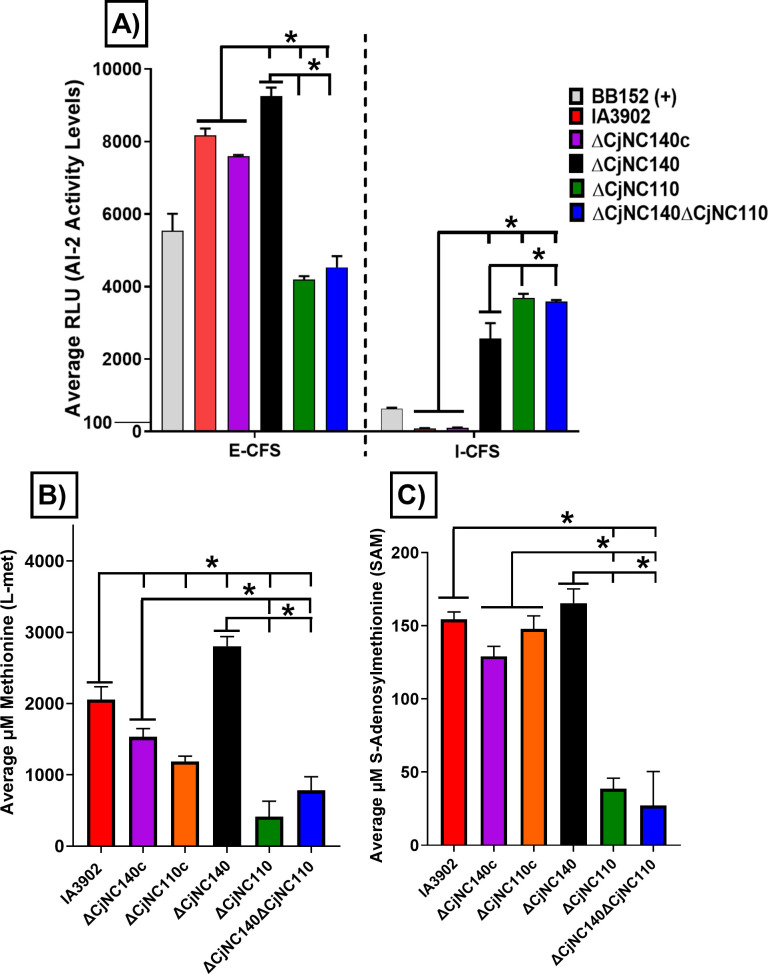
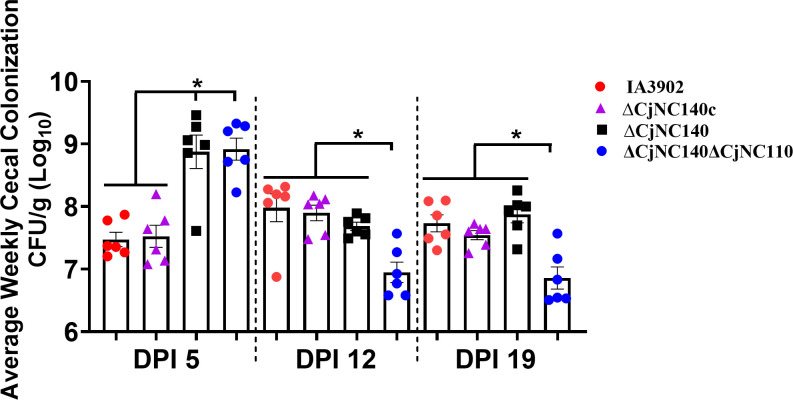
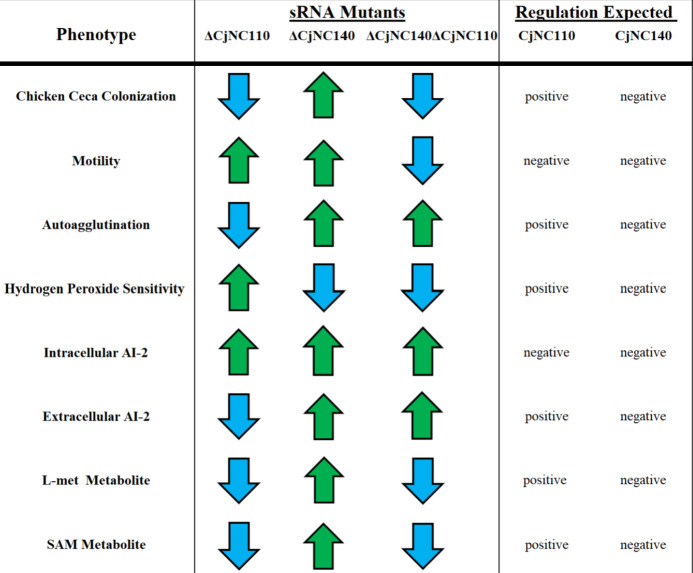
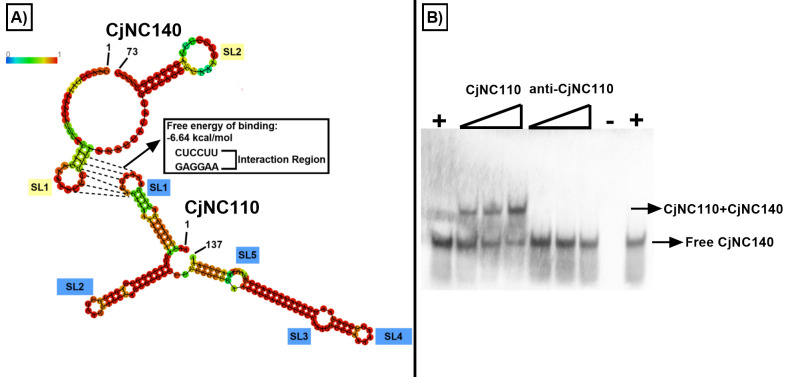
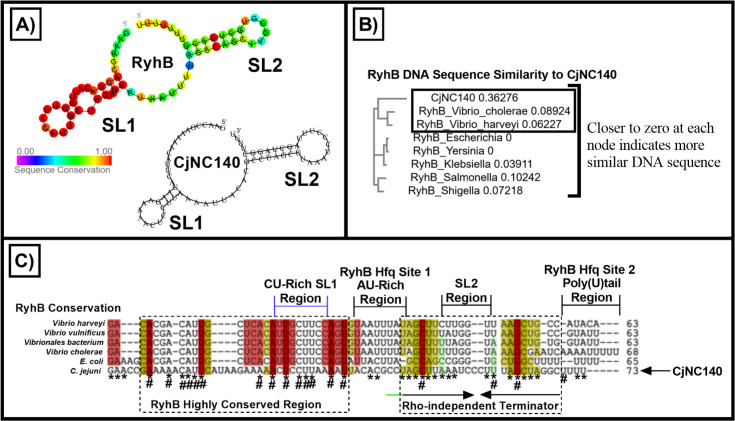
Small non-coding RNAs (sRNAs) are important players in modulating gene expression in bacterial pathogens, but their functions are largely undetermined in Campylobacter jejuni, an important cause of foodborne gastroenteritis in humans. In this study, we elucidated the functions of sRNA CjNC140 and its interaction with CjNC110, a previously characterized sRNA involved in the regulation of several virulence phenotypes of C. jejuni. Inactivation of CjNC140 increased motility, autoagglutination, L-methionine concentration, autoinducer-2 production, hydrogen peroxide resistance, and early chicken colonization, indicating a primarily inhibitory role of CjNC140 for these phenotypes. Apart from motility, all these effects directly contrasted the previously demonstrated positive regulation by CjNC110, suggesting that CjNC110 and CjNC140 operate in an opposite manner to modulate physiologic processes in C. jejuni. RNAseq and northern blotting further demonstrated that expression of CjNC140 increased in the absence of CjNC110, while expression of CjNC110 decreased in the absence of CjNC140, suggesting a possibility of their direct interaction. Indeed, electrophoretic mobility shift assay demonstrated a direct binding between the two sRNAs via GA- (CjNC110) and CU- (CjNC140) rich stem-loops. Additionally, RNAseq and follow-up experiments identified that CjNC140 positively regulates p19, which encodes a key iron uptake transporter in Campylobacter. Furthermore, computational analysis revealed both CjNC140 and CjNC110 are highly conserved in C. jejuni, and the predicted secondary structures support CjNC140 as a functional homolog of the iron regulatory sRNA, RyhB. These findings establish CjNC140 and CjNC110 as a key checks-and- balances mechanism in maintaining homeostasis of gene expression and optimizing phenotypes critical for C. jejuni pathobiology. IMPORTANCE Gene regulation is critical to all aspects of pathogenesis of bacterial disease, and small non-coding RNAs (sRNAs) represent a new frontier in gene regulation of bacteria. In Campylobacter jejuni, the role of sRNAs remains largely unexplored. Here, we investigate the role of two highly conserved sRNAs, CjNC110 and CjNC140, and demonstrate that CjNC140 displays a primarily inhibitory role in contrast to a primarily activating role for CjNC110 for several key virulence-associated phenotypes. Our results also revealed that the sRNA regulatory pathway is intertwined with the iron uptake system, another virulence mechanism critical for in vivo colonization. These findings open a new direction for understanding C. jejuni pathobiology and identify potential targets for intervention for this major foodborne pathogen.
Keywords: Campylobacter; iron homeostasis; post-transcriptional regulation; small non-coding RNAs; sponge RNAs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Small RNA CjNC110 regulates the activated methyl cycle to enable optimal chicken colonization by Campylobacter jejuni.mSphere. 2025 Jan 28;10(1):e0083224. doi: 10.1128/msphere.00832-24. Epub 2025 Jan 8. mSphere. 2025. PMID: 39772717 Free PMC article.
-
Small Noncoding RNA CjNC110 Influences Motility, Autoagglutination, AI-2 Localization, Hydrogen Peroxide Sensitivity, and Chicken Colonization in Campylobacter jejuni.Infect Immun. 2020 Jun 22;88(7):e00245-20. doi: 10.1128/IAI.00245-20. Print 2020 Jun 22. Infect Immun. 2020. PMID: 32366573 Free PMC article.
-
Posttranscriptional Regulation in Response to Different Environmental Stresses in Campylobacter jejuni.Microbiol Spectr. 2022 Jun 29;10(3):e0020322. doi: 10.1128/spectrum.00203-22. Epub 2022 Jun 9. Microbiol Spectr. 2022. PMID: 35678555 Free PMC article.
-
Interplay between iron homeostasis and virulence: Fur and RyhB as major regulators of bacterial pathogenicity.Vet Microbiol. 2015 Aug 31;179(1-2):2-14. doi: 10.1016/j.vetmic.2015.03.024. Epub 2015 Apr 8. Vet Microbiol. 2015. PMID: 25888312 Review.
-
Polyphosphate and associated enzymes as global regulators of stress response and virulence in Campylobacter jejuni.World J Gastroenterol. 2016 Sep 7;22(33):7402-14. doi: 10.3748/wjg.v22.i33.7402. World J Gastroenterol. 2016. PMID: 27672264 Free PMC article. Review.
Cited by
-
Small RNA CjNC110 regulates the activated methyl cycle to enable optimal chicken colonization by Campylobacter jejuni.mSphere. 2025 Jan 28;10(1):e0083224. doi: 10.1128/msphere.00832-24. Epub 2025 Jan 8. mSphere. 2025. PMID: 39772717 Free PMC article.
References
-
- Sahin O, Plummer PJ, Jordan DM, Sulaj K, Pereira S, Robbe-Austerman S, Wang L, Yaeger MJ, Hoffman LJ, Zhang Q. 2008. Emergence of a tetracycline-resistant Campylobacter jejuni clone associated with outbreaks of ovine abortion in the United States. J Clin Microbiol 46:1663–1671. doi:10.1128/JCM.00031-08 - DOI - PMC - PubMed
-
- Sahin O, Fitzgerald C, Stroika S, Zhao S, Sippy RJ, Kwan P, Plummer PJ, Han J, Yaeger MJ, Zhang Q. 2011. Molecular evidence for zoonotic transmission of an emergent, highly pathogenic Campylobacter jejuni clone in the United States. J Clin Microbiol 50:680–687. doi:10.1128/JCM.06167-11 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
