

Differential Modulation of GABAergic and Glutamatergic Neurons in the Ventral Pallidum by GABA and Neuropeptides
- PMID: 37414552
- PMCID: PMC10348443
- DOI: 10.1523/ENEURO.0404-22.2023
Differential Modulation of GABAergic and Glutamatergic Neurons in the Ventral Pallidum by GABA and Neuropeptides
Abstract
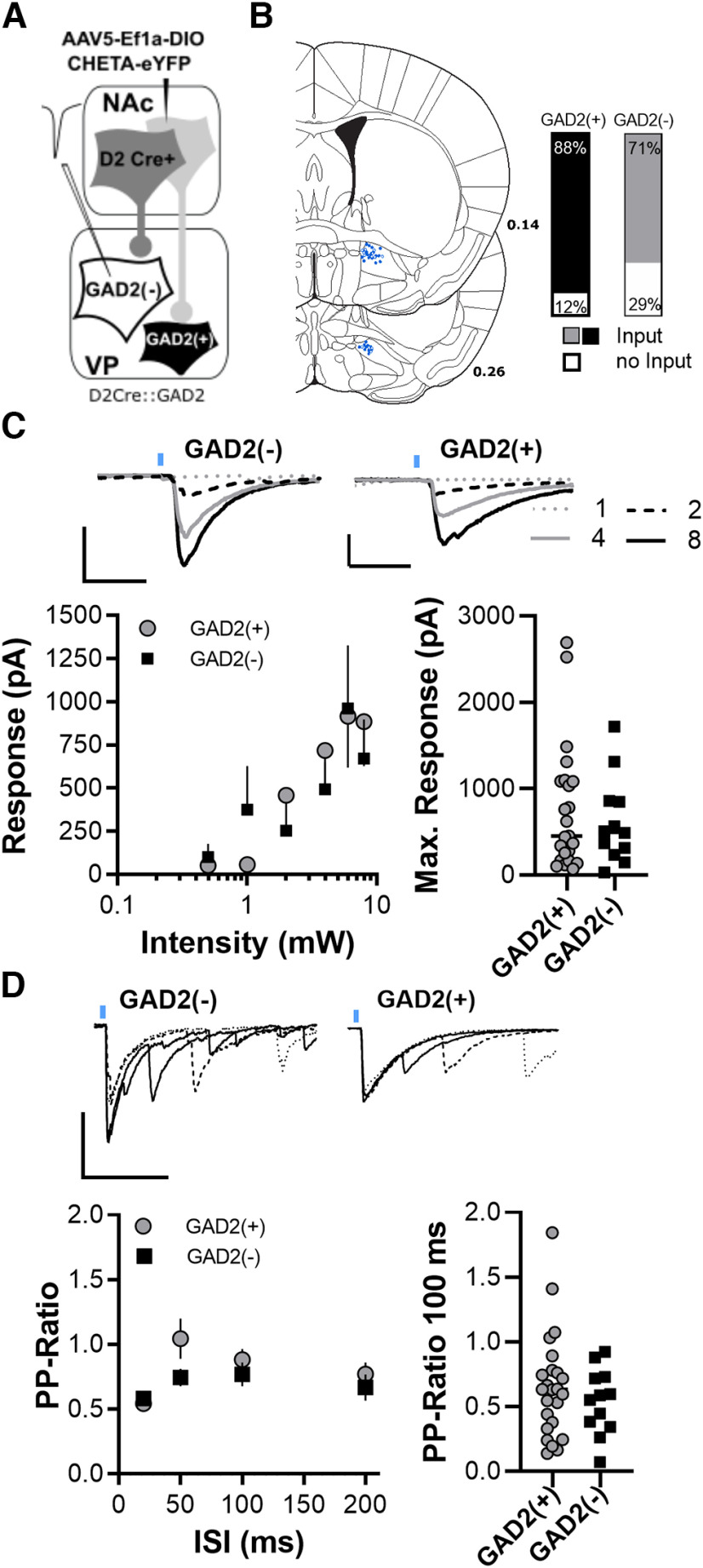
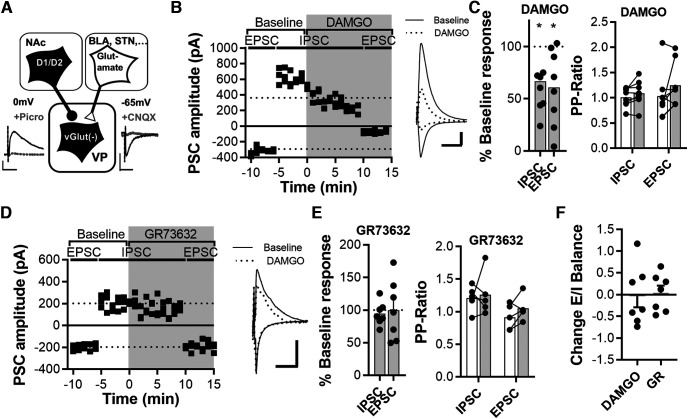
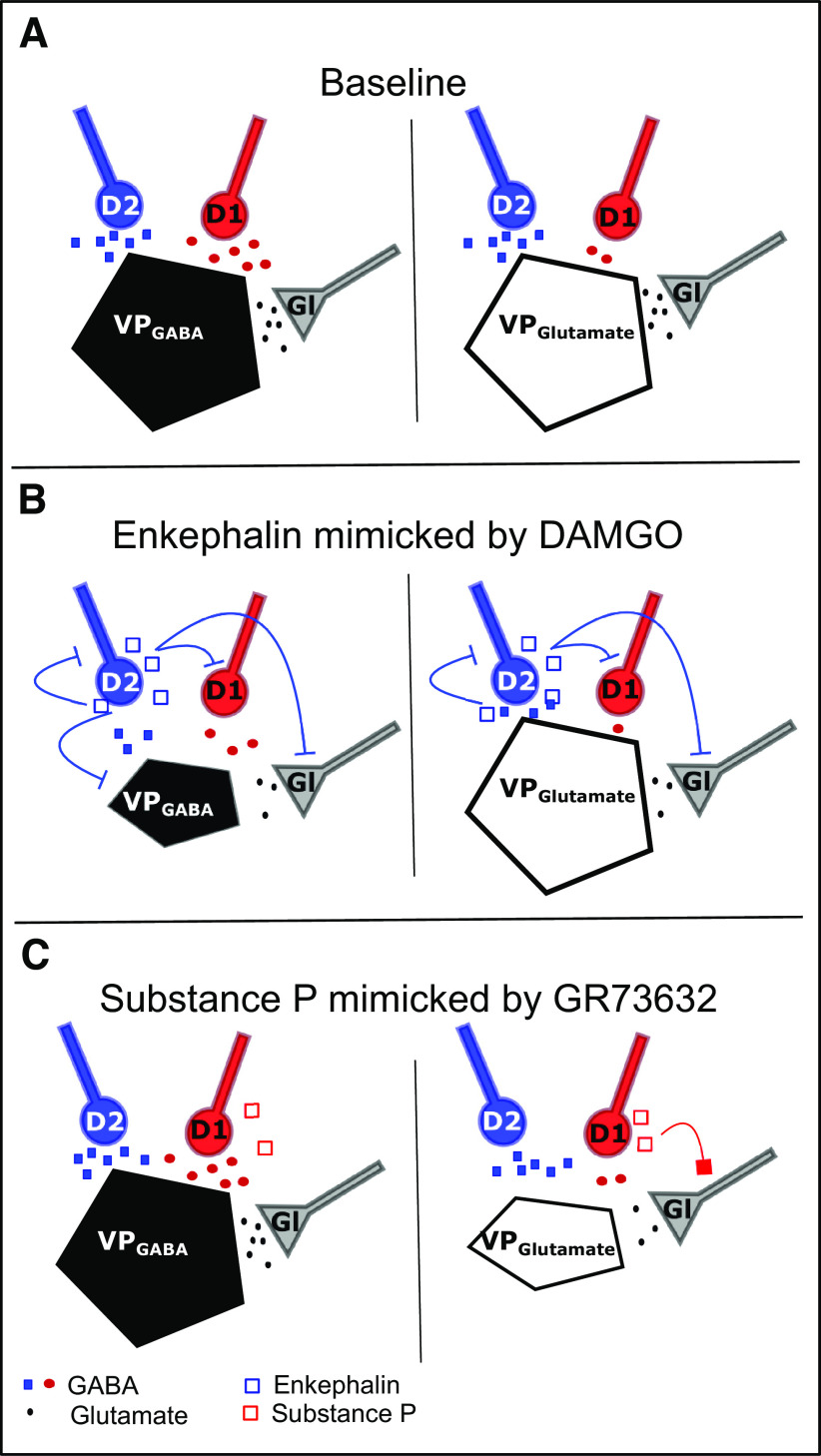
The ventral pallidum (VP) is an integral locus in the reward circuitry and a major target of GABAergic innervation of both D1-medium spiny neurons (MSNs) and D2-MSNs from the nucleus accumbens. The VP contains populations of GABAergic [VPGABA, GAD2(+), or VGluT(-)] and glutamatergic [VPGlutamate, GAD2(-), or VGluT(+)] cells that facilitate positive reinforcement and behavioral avoidance, respectively. MSN efferents to the VP exert opponent control over behavioral reinforcement with activation of D1-MSN afferents promoting and D2-MSN afferents inhibiting reward seeking. How this afferent-specific and cell type-specific control of reward seeking is integrated remains largely unknown. In addition to GABA, D1-MSNs corelease substance P to stimulate neurokinin 1 receptors (NK1Rs) and D2-MSNs corelease enkephalin to activate μ-opioid receptors (MORs) and δ-opioid receptors. These neuropeptides act in the VP to alter appetitive behavior and reward seeking. Using a combination of optogenetics and patch-clamp electrophysiology in mice, we found that GAD2(-) cells receive weaker GABA input from D1-MSN, but GAD2(+) cells receive comparable GABAergic input from both afferent types. Pharmacological activation of MORs induced an equally strong presynaptic inhibition of GABA and glutamate transmission on both cell types. Interestingly, MOR activation hyperpolarized VPGABA but not VGluT(+). NK1R activation inhibited glutamatergic transmission only on VGluT(+) cells. Our results indicate that the afferent-specific release of GABA and neuropeptides from D1-MSNs and D2-MSNs can differentially influence VP neuronal subtypes.
Keywords: GABA; medium spiny neurons; neuropeptides; optogenetics; ventral pallidumg; whole cell patch clamp.
Copyright © 2023 Neuhofer and Kalivas.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous