

Diacylglycerol lipase alpha promotes hepatocellular carcinoma progression and induces lenvatinib resistance by enhancing YAP activity
- PMID: 37414748
- PMCID: PMC10325985
- DOI: 10.1038/s41419-023-05919-5
Diacylglycerol lipase alpha promotes hepatocellular carcinoma progression and induces lenvatinib resistance by enhancing YAP activity
Abstract
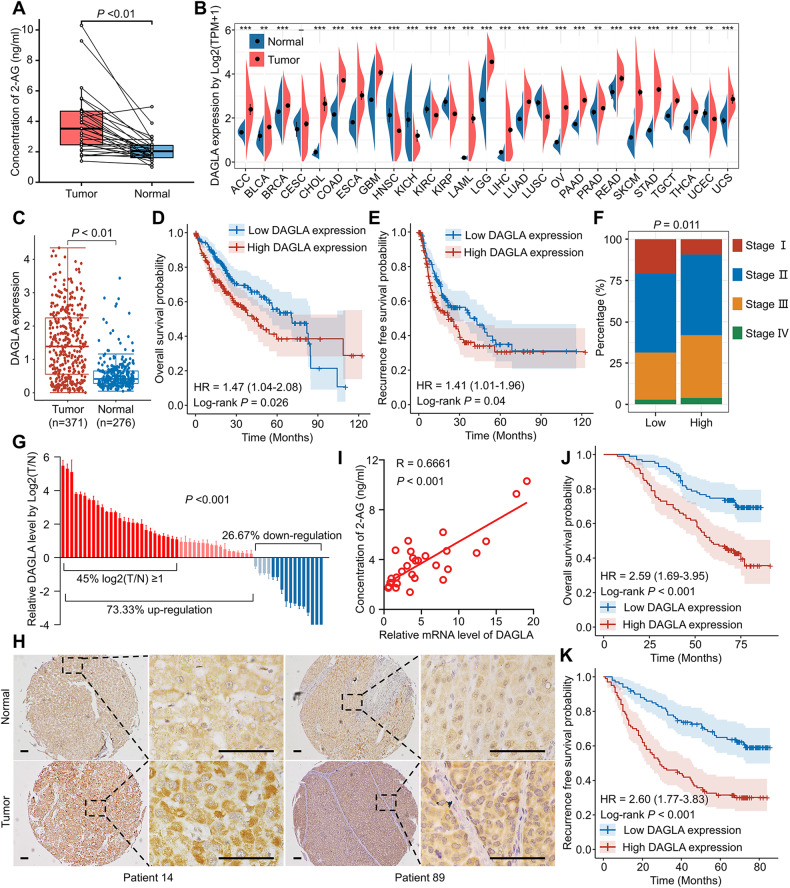
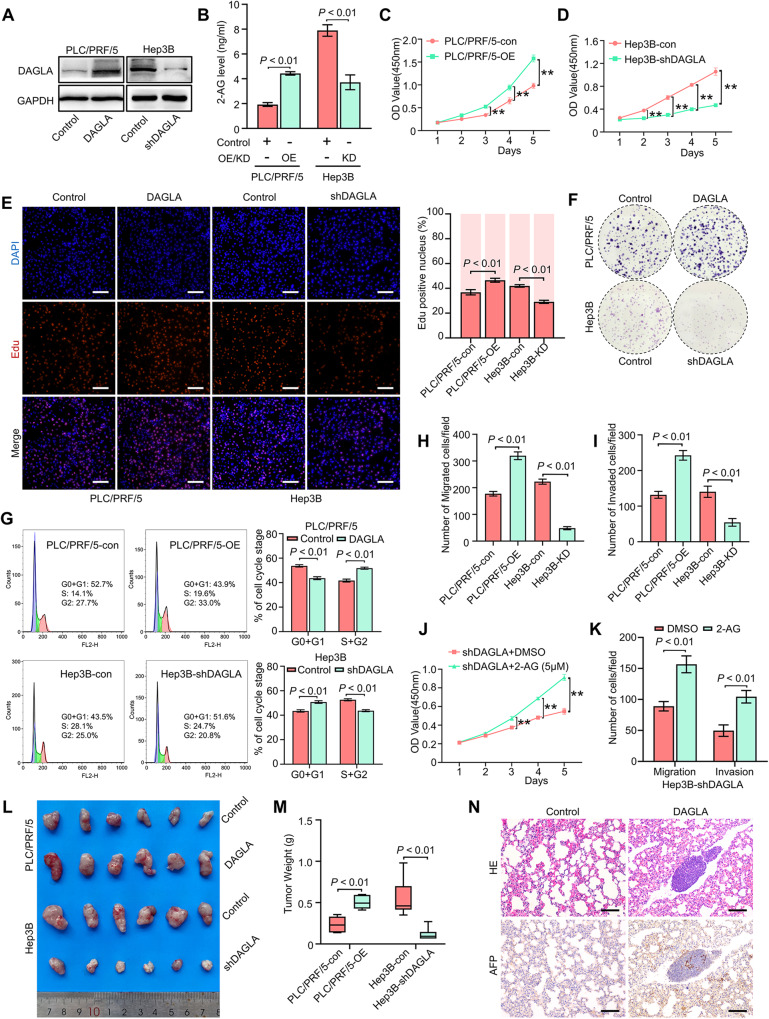
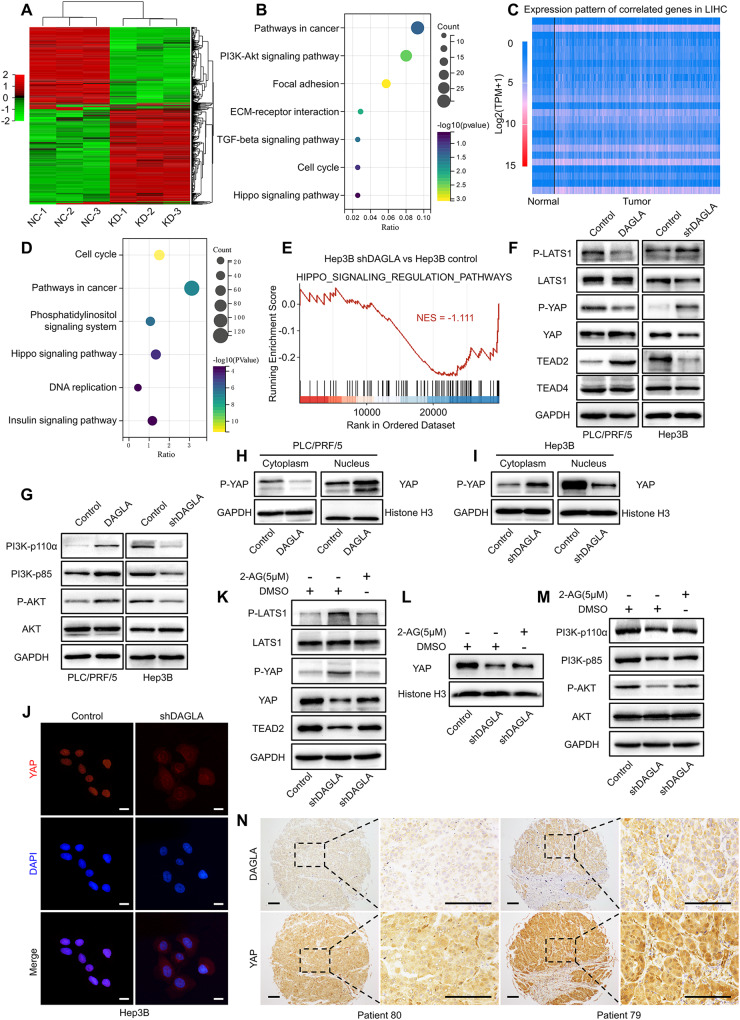
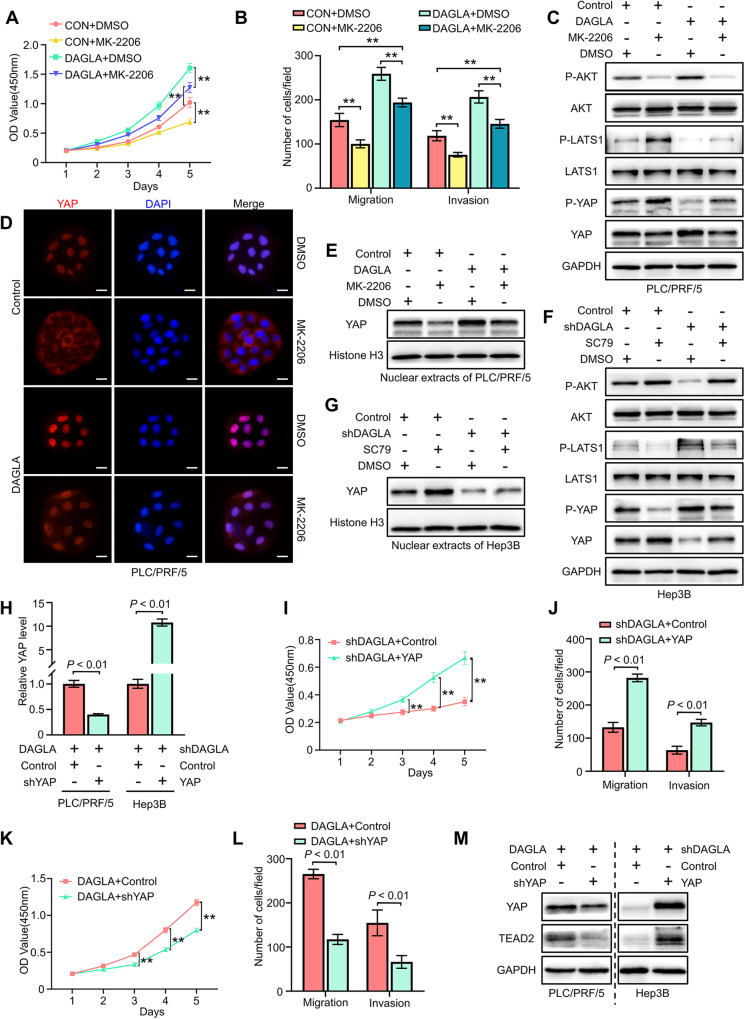
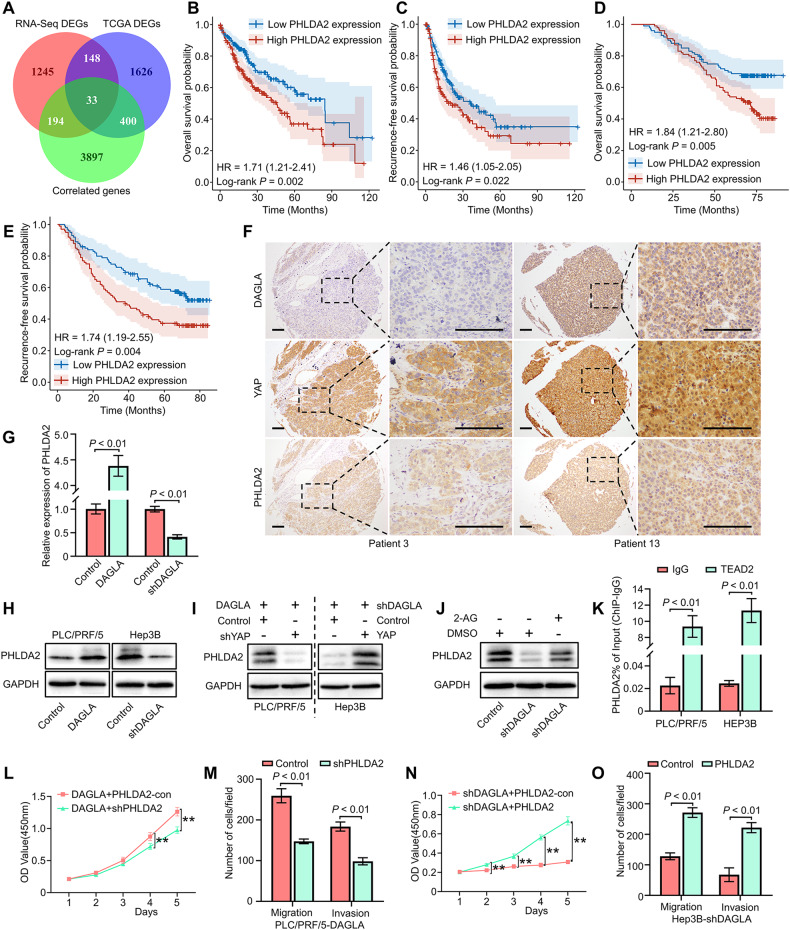
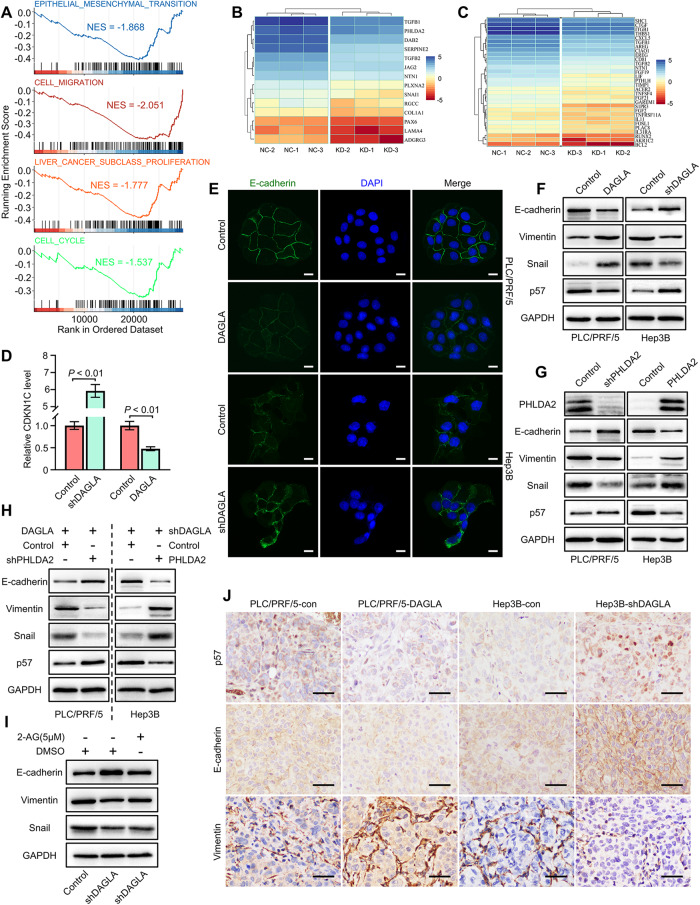
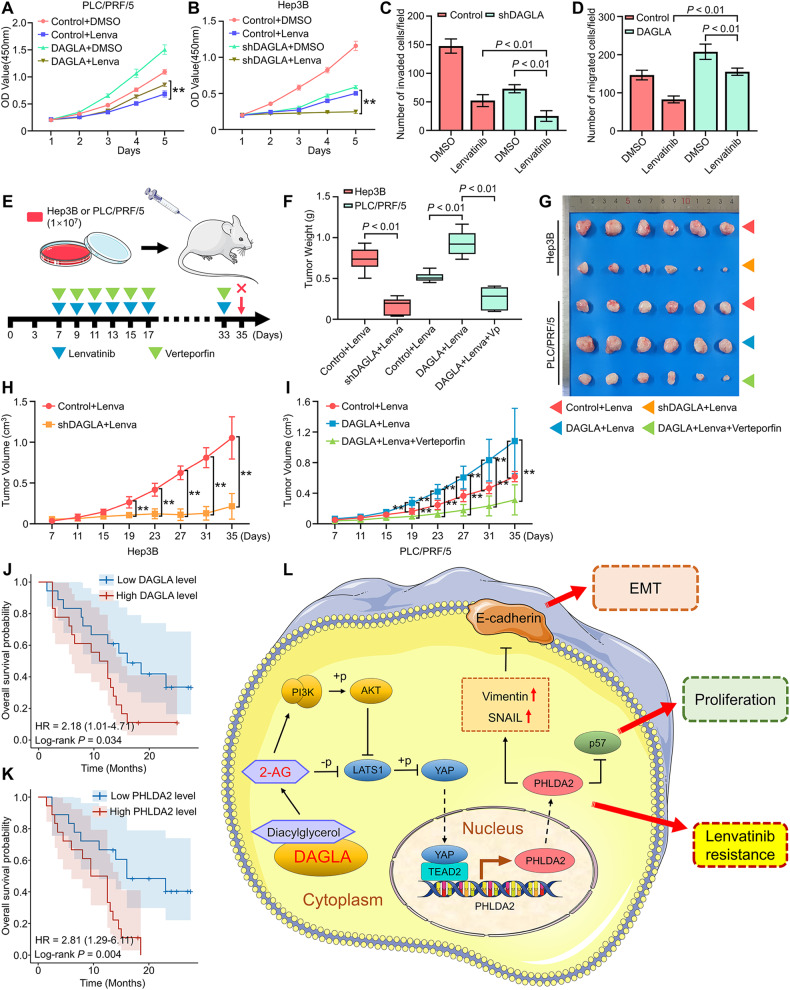
As an important hydrolytic enzyme that yields 2-AG and free fatty acids, diacylglycerol lipase alpha (DAGLA) is involved in exacerbating malignant phenotypes and cancer progression, but the role of the DAGLA/2-AG axis in HCC progression remains unclear. Here, we found that the upregulation of components of the DAGLA/2-AG axis in HCC samples is correlated with tumour stage and patient prognosis. In vitro and in vivo experiments demonstrated that the DAGLA/2-AG axis promoted HCC progression by regulating cell proliferation, invasion and metastasis. Mechanistically, the DAGLA/2AG axis significantly inhibited LATS1 and YAP phosphorylation, promoted YAP nuclear translocation and activity, and ultimately led to TEAD2 upregulation and increased PHLDA2 expression, which could be enhanced by DAGLA/2AG-induced activation of the PI3K/AKT pathway. More importantly, DAGLA induced resistance to lenvatinib therapy during HCC treatment. Our study demonstrates that inhibiting the DAGLA/2-AG axis could be a novel therapeutic strategy to inhibit HCC progression and enhance the therapeutic effects of TKIs, which warrant further clinical studies.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
