

Intronic small nucleolar RNAs regulate host gene splicing through base pairing with their adjacent intronic sequences
- PMID: 37415181
- PMCID: PMC10324135
- DOI: 10.1186/s13059-023-03002-y
Intronic small nucleolar RNAs regulate host gene splicing through base pairing with their adjacent intronic sequences
Abstract
Background: Small nucleolar RNAs (snoRNAs) are abundant noncoding RNAs best known for their involvement in ribosomal RNA maturation. In mammals, most expressed snoRNAs are embedded in introns of longer genes and produced through transcription and splicing of their host. Intronic snoRNAs were long viewed as inert passengers with little effect on host expression. However, a recent study reported a snoRNA influencing the splicing and ultimate output of its host gene. Overall, the general contribution of intronic snoRNAs to host expression remains unclear.
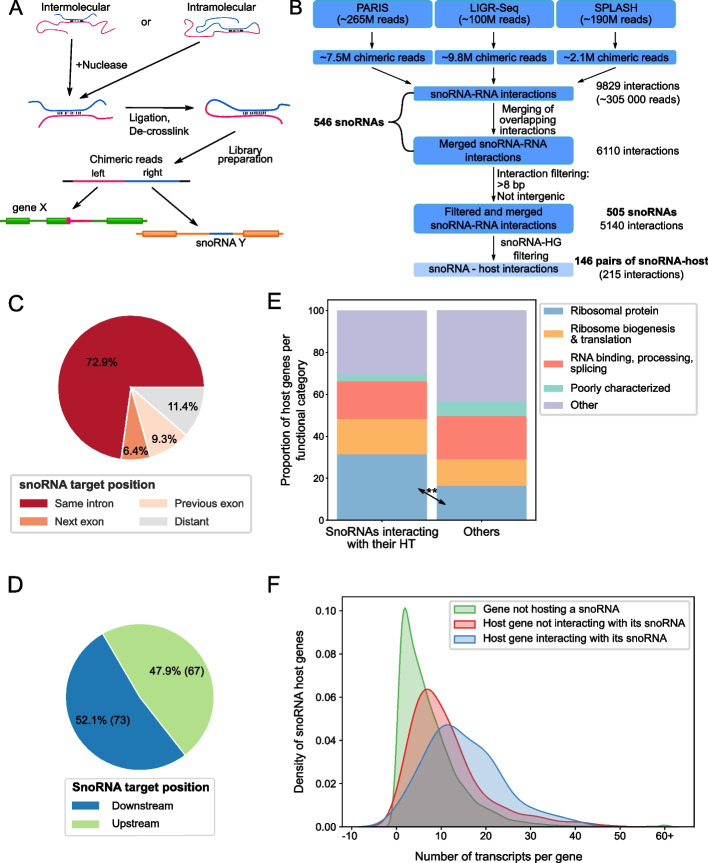
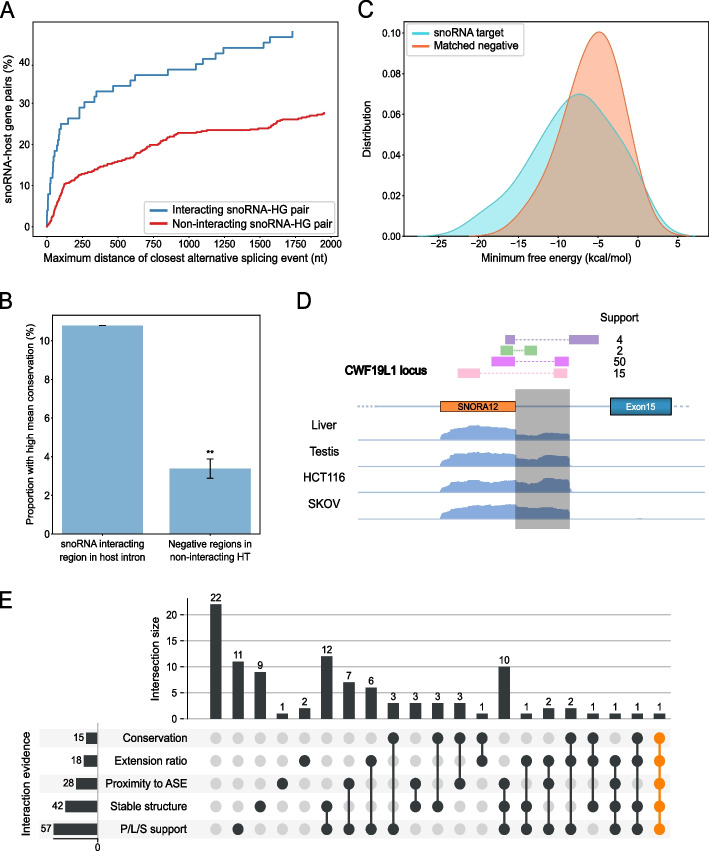
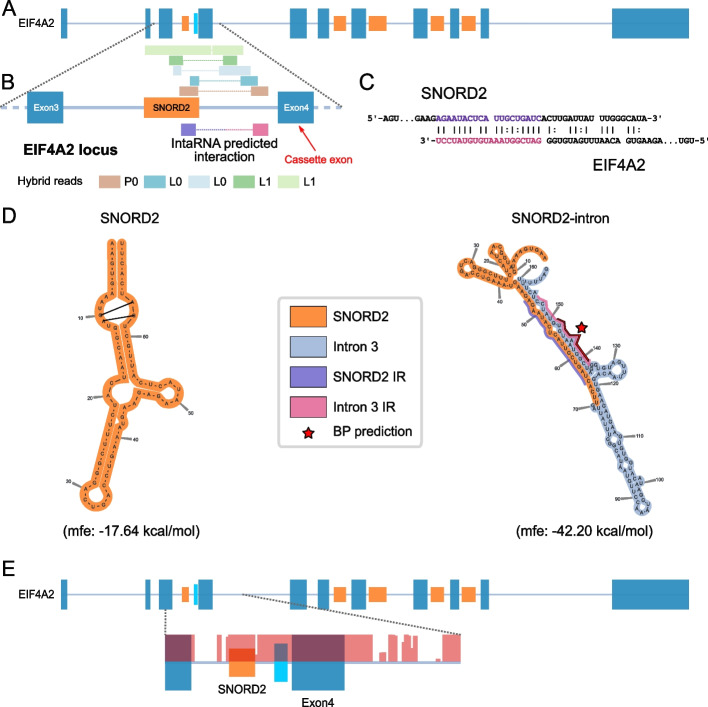
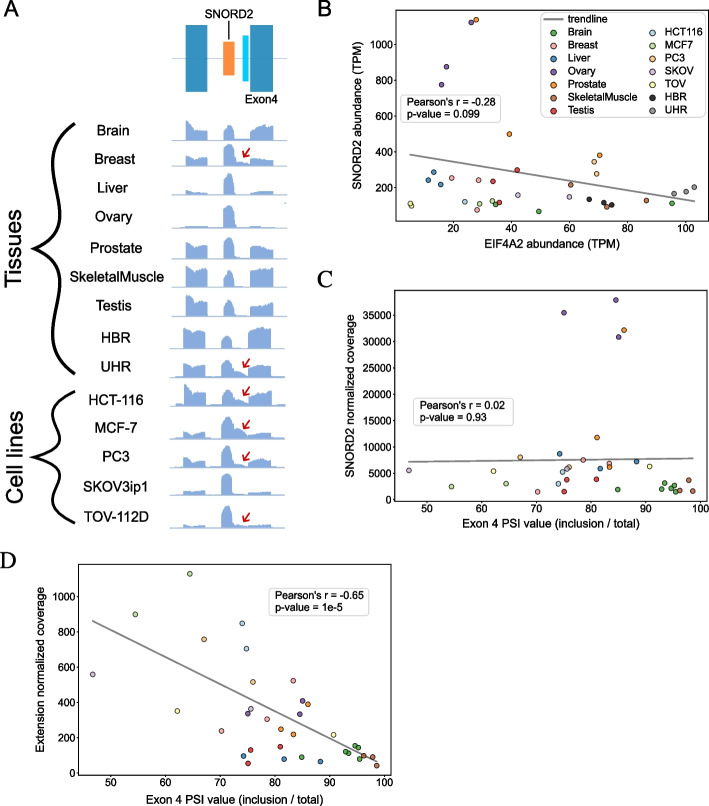
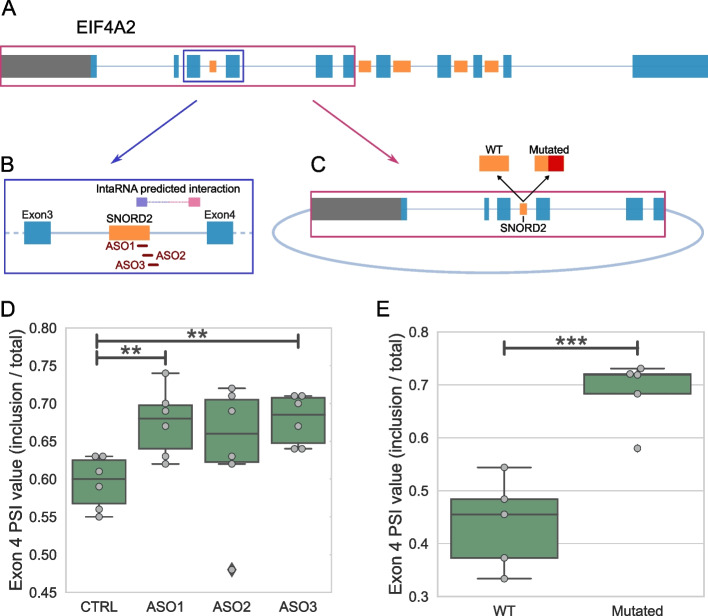
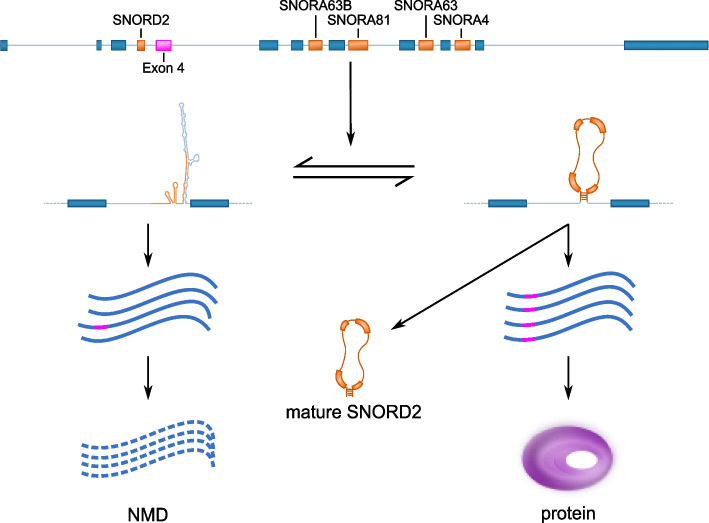
Results: Computational analysis of large-scale human RNA-RNA interaction datasets indicates that 30% of detected snoRNAs interact with their host transcripts. Many snoRNA-host duplexes are located near alternatively spliced exons and display high sequence conservation suggesting a possible role in splicing regulation. The study of the model SNORD2-EIF4A2 duplex indicates that the snoRNA interaction with the host intronic sequence conceals the branch point leading to decreased inclusion of the adjacent alternative exon. Extended SNORD2 sequence containing the interacting intronic region accumulates in sequencing datasets in a cell-type-specific manner. Antisense oligonucleotides and mutations that disrupt the formation of the snoRNA-intron structure promote the splicing of the alternative exon, shifting the EIF4A2 transcript ratio away from nonsense-mediated decay.
Conclusions: Many snoRNAs form RNA duplexes near alternative exons of their host transcripts, placing them in optimal positions to control host output as shown for the SNORD2-EIF4A2 model system. Overall, our study supports a more widespread role for intronic snoRNAs in the regulation of their host transcript maturation.
Keywords: Alternative splicing regulation; Cis-regulation; Intron; Nonsense-mediated decay; RNA secondary structure; RNA-RNA interactions; SnoRNA; SnoRNA/host gene relationship.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
