

Insights into the Structure of Comirnaty Covid-19 Vaccine: A Theory on Soft, Partially Bilayer-Covered Nanoparticles with Hydrogen Bond-Stabilized mRNA-Lipid Complexes
- PMID: 37417667
- PMCID: PMC10373524
- DOI: 10.1021/acsnano.2c11904
Insights into the Structure of Comirnaty Covid-19 Vaccine: A Theory on Soft, Partially Bilayer-Covered Nanoparticles with Hydrogen Bond-Stabilized mRNA-Lipid Complexes
Abstract
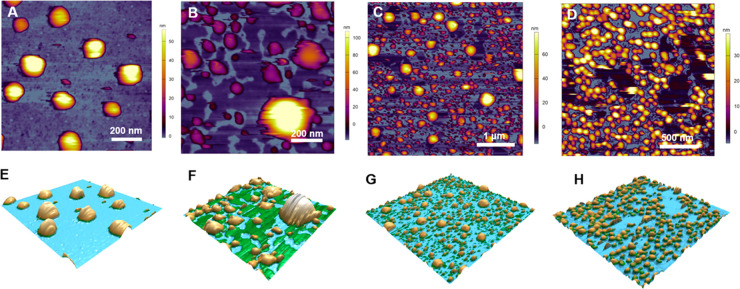
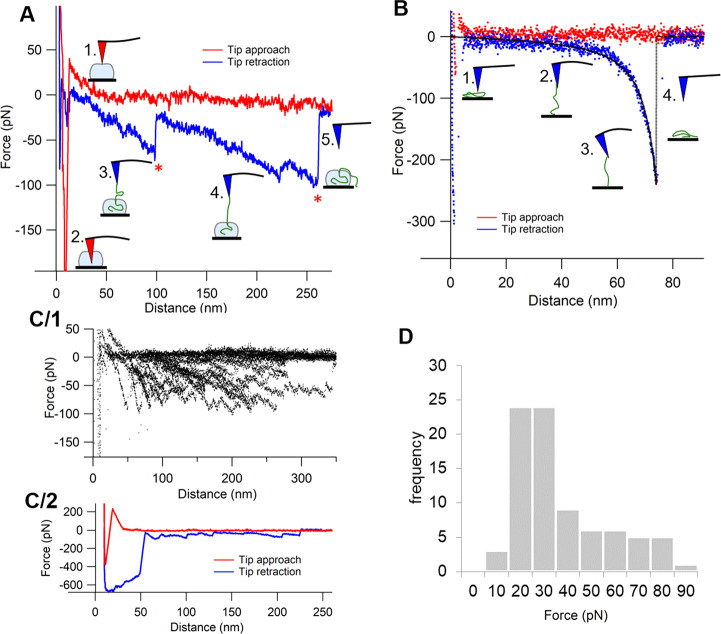
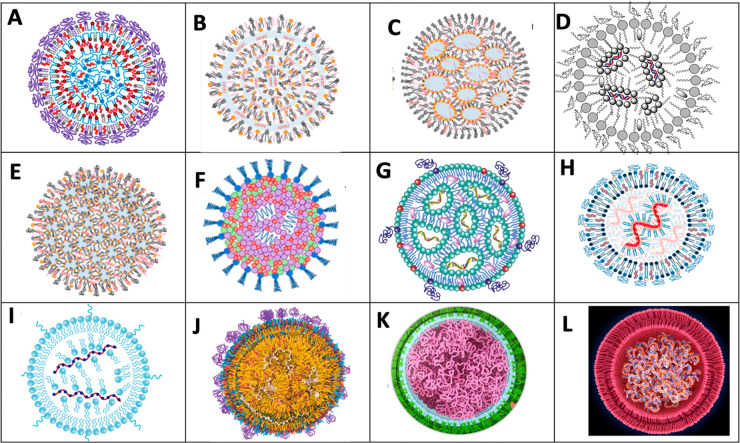
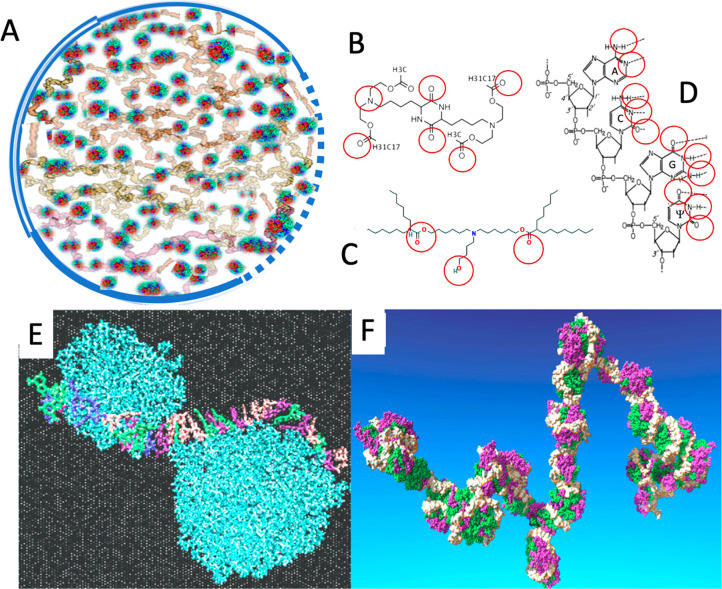
Despite the worldwide success of mRNA-LNP Covid-19 vaccines, the nanoscale structures of these formulations are still poorly understood. To fill this gap, we used a combination of atomic force microscopy (AFM), dynamic light scattering (DLS), transmission electron microscopy (TEM), cryogenic transmission electron microscopy (cryo-TEM), and the determination of the intra-LNP pH gradient to analyze the nanoparticles (NPs) in BNT162b2 (Comirnaty), comparing it with the well-characterized PEGylated liposomal doxorubicin (Doxil). Comirnaty NPs had similar size and envelope lipid composition to Doxil; however, unlike Doxil liposomes, wherein the stable ammonium and pH gradient enables accumulation of 14C-methylamine in the intraliposomal aqueous phase, Comirnaty LNPs lack such pH gradient in spite of the fact that the pH 4, at which LNPs are prepared, is raised to pH 7.2 after loading of the mRNA. Mechanical manipulation of Comirnaty NPs with AFM revealed soft, compliant structures. The sawtooth-like force transitions seen during cantilever retraction imply that molecular strands, corresponding to mRNA, can be pulled out of NPs, and the process is accompanied by stepwise rupture of mRNA-lipid bonds. Unlike Doxil, cryo-TEM of Comirnaty NPs revealed a granular, solid core enclosed by mono- and bilipid layers. Negative staining TEM shows 2-5 nm electron-dense spots in the LNP's interior that are aligned into strings, semicircles, or labyrinth-like networks, which may imply cross-link-stabilized RNA fragments. The neutral intra-LNP core questions the dominance of ionic interactions holding together this scaffold, raising the possibility of hydrogen bonding between mRNA and the lipids. Such interaction, described previously for another mRNA/lipid complex, is consistent with the steric structure of the ionizable lipid in Comirnaty, ALC-0315, displaying free ═O and -OH groups. It is hypothesized that the latter groups can get into steric positions that enable hydrogen bonding with the nitrogenous bases in the mRNA. These structural features of mRNA-LNP may be important for the vaccine's activities in vivo.
Keywords: Doxil liposomes; SARS-CoV-2; atomic force microscopy; cryo-electron microscopy; dynamic light scattering; lipid nanoparticles; phospholipid membranes.
Conflict of interest statement
The authors declare no competing financial interest.
Figures


Similar articles
-
Mesoscopic Structure of Lipid Nanoparticle Formulations for mRNA Drug Delivery: Comirnaty and Drug-Free Dispersions.ACS Nano. 2024 Apr 2;18(13):9746-9764. doi: 10.1021/acsnano.4c02610. Epub 2024 Mar 21. ACS Nano. 2024. PMID: 38514237
-
Calculating Apparent pKa Values of Ionizable Lipids in Lipid Nanoparticles.Mol Pharm. 2025 Jan 6;22(1):588-593. doi: 10.1021/acs.molpharmaceut.4c00426. Epub 2024 Dec 10. Mol Pharm. 2025. PMID: 39655829 Free PMC article.
-
Anti-PEG antibodies before and after a first dose of Comirnaty® (mRNA-LNP-based SARS-CoV-2 vaccine).J Control Release. 2023 Feb;354:316-322. doi: 10.1016/j.jconrel.2022.12.039. Epub 2023 Jan 13. J Control Release. 2023. PMID: 36549393 Free PMC article.
-
Chemistry of Lipid Nanoparticles for RNA Delivery.Acc Chem Res. 2022 Jan 4;55(1):2-12. doi: 10.1021/acs.accounts.1c00544. Epub 2021 Dec 1. Acc Chem Res. 2022. PMID: 34850635 Review.
-
Rational design and combinatorial chemistry of ionizable lipids for RNA delivery.J Mater Chem B. 2023 Jul 19;11(28):6527-6539. doi: 10.1039/d3tb00649b. J Mater Chem B. 2023. PMID: 37345430 Review.
Cited by
-
Nanoscience in Action: Unveiling Emerging Trends in Materials and Applications.ACS Omega. 2025 Feb 17;10(8):7530-7548. doi: 10.1021/acsomega.4c10929. eCollection 2025 Mar 4. ACS Omega. 2025. PMID: 40060806 Free PMC article. Review.
-
Lipid Nanoparticle (LNP) Delivery Carrier-Assisted Targeted Controlled Release mRNA Vaccines in Tumor Immunity.Vaccines (Basel). 2024 Feb 12;12(2):186. doi: 10.3390/vaccines12020186. Vaccines (Basel). 2024. PMID: 38400169 Free PMC article. Review.
-
Advances in locally administered nucleic acid therapeutics.Bioact Mater. 2025 Mar 10;49:218-254. doi: 10.1016/j.bioactmat.2025.02.043. eCollection 2025 Jul. Bioact Mater. 2025. PMID: 40144794 Free PMC article. Review.
-
Mechanisms and Barriers in Nanomedicine: Progress in the Field and Future Directions.ACS Nano. 2024 Jun 4;18(22):13983-13999. doi: 10.1021/acsnano.4c00182. Epub 2024 May 20. ACS Nano. 2024. PMID: 38767983 Free PMC article.
-
Expanded Spectrum and Increased Incidence of Adverse Events Linked to COVID-19 Genetic Vaccines: New Concepts on Prophylactic Immuno-Gene Therapy, Iatrogenic Orphan Disease, and Platform-Inherent Challenges.Pharmaceutics. 2025 Mar 31;17(4):450. doi: 10.3390/pharmaceutics17040450. Pharmaceutics. 2025. PMID: 40284445 Free PMC article. Review.
References
-
- Sahin U.; Muik A.; Derhovanessian E.; Vogler I.; Kranz L. M.; Vormehr M.; Baum A.; Pascal K.; Quandt J.; Maurus D.; Brachtendorf S.; Lorks V.; Sikorski J.; Hilker R.; Becker D.; Eller A. K.; Grutzner J.; Boesler C.; Rosenbaum C.; Kuhnle M. C.; Luxemburger U.; Kemmer-Bruck A.; Langer D.; Bexon M.; Bolte S.; Kariko K.; Palanche T.; Fischer B.; Schultz A.; Shi P. Y.; Fontes-Garfias C.; Perez J. L.; Swanson K. A.; Loschko J.; Scully I. L.; Cutler M.; Kalina W.; Kyratsous C. A.; Cooper D.; Dormitzer P. R.; Jansen K. U.; Tureci O. COVID-19 vaccine BNT162b1 elicits human antibody and T(H) 1 T cell responses. Nature 2020, 586 (7830), 594–599. 10.1038/s41586-020-2814-7. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
