

Generating high-fidelity cochlear organoids from human pluripotent stem cells
- PMID: 37419105
- PMCID: PMC10695300
- DOI: 10.1016/j.stem.2023.06.006
Generating high-fidelity cochlear organoids from human pluripotent stem cells
Abstract
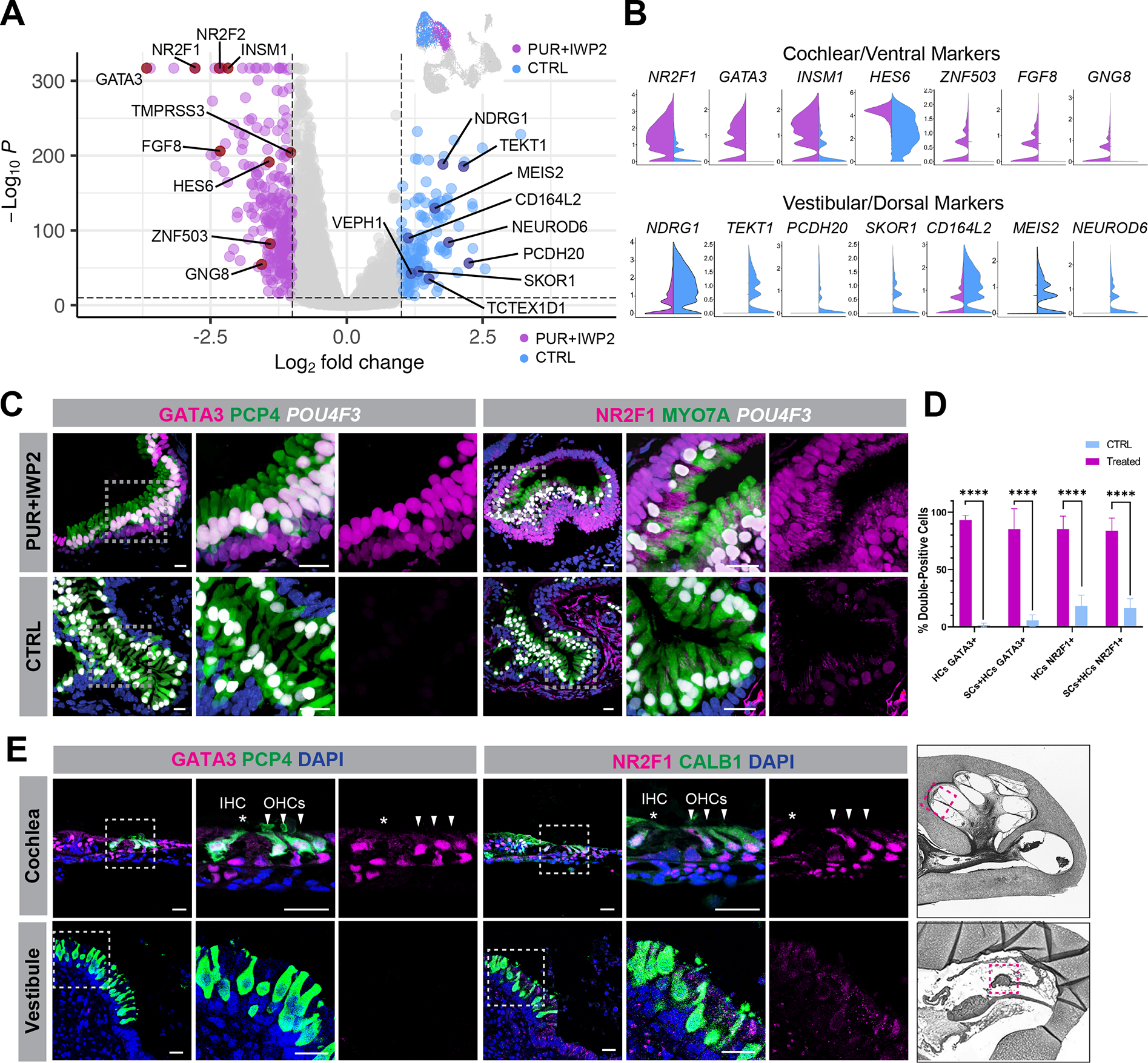
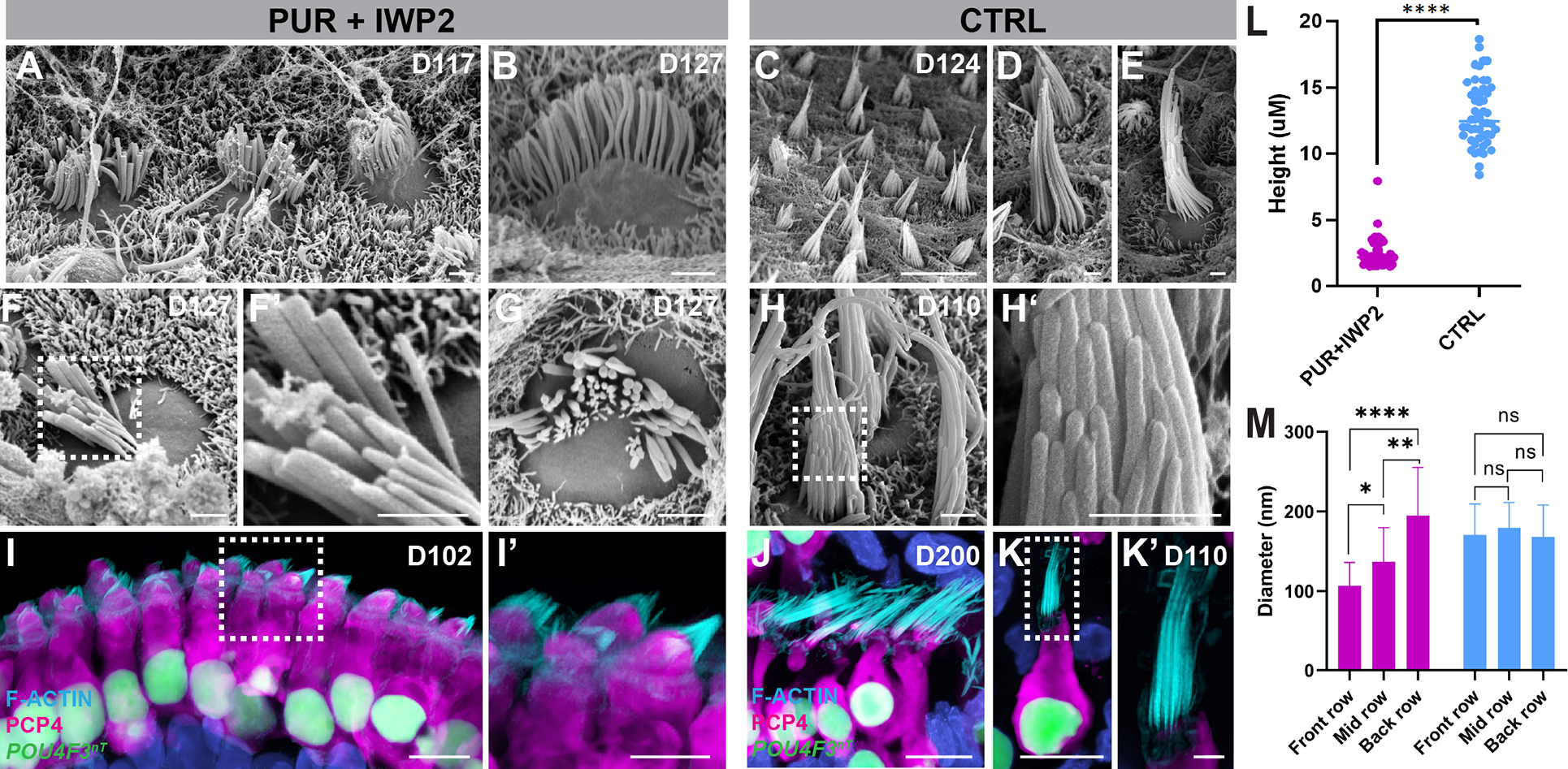
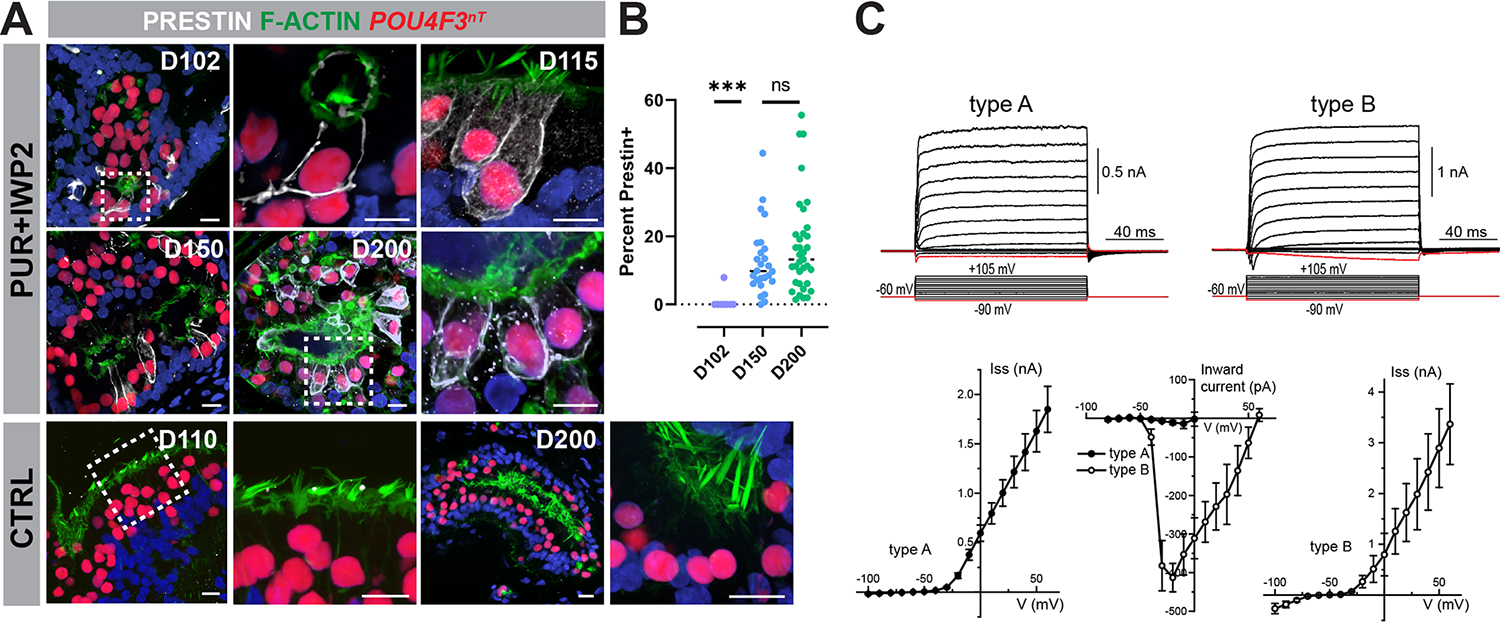
Mechanosensitive hair cells in the cochlea are responsible for hearing but are vulnerable to damage by genetic mutations and environmental insults. The paucity of human cochlear tissues makes it difficult to study cochlear hair cells. Organoids offer a compelling platform to study scarce tissues in vitro; however, derivation of cochlear cell types has proven non-trivial. Here, using 3D cultures of human pluripotent stem cells, we sought to replicate key differentiation cues of cochlear specification. We found that timed modulations of Sonic Hedgehog and WNT signaling promote ventral gene expression in otic progenitors. Ventralized otic progenitors subsequently give rise to elaborately patterned epithelia containing hair cells with morphology, marker expression, and functional properties consistent with both outer and inner hair cells in the cochlea. These results suggest that early morphogenic cues are sufficient to drive cochlear induction and establish an unprecedented system to model the human auditory organ.
Keywords: auditory; cochlea; hair cell; human; inner ear; organoid; pluripotent stem cell; scRNA-seq.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.T.M. and E.H. are the inventors on a PCT patent application titled “Methods of generating human cochlear hair cells.” The other authors declare no competing interests.
Figures
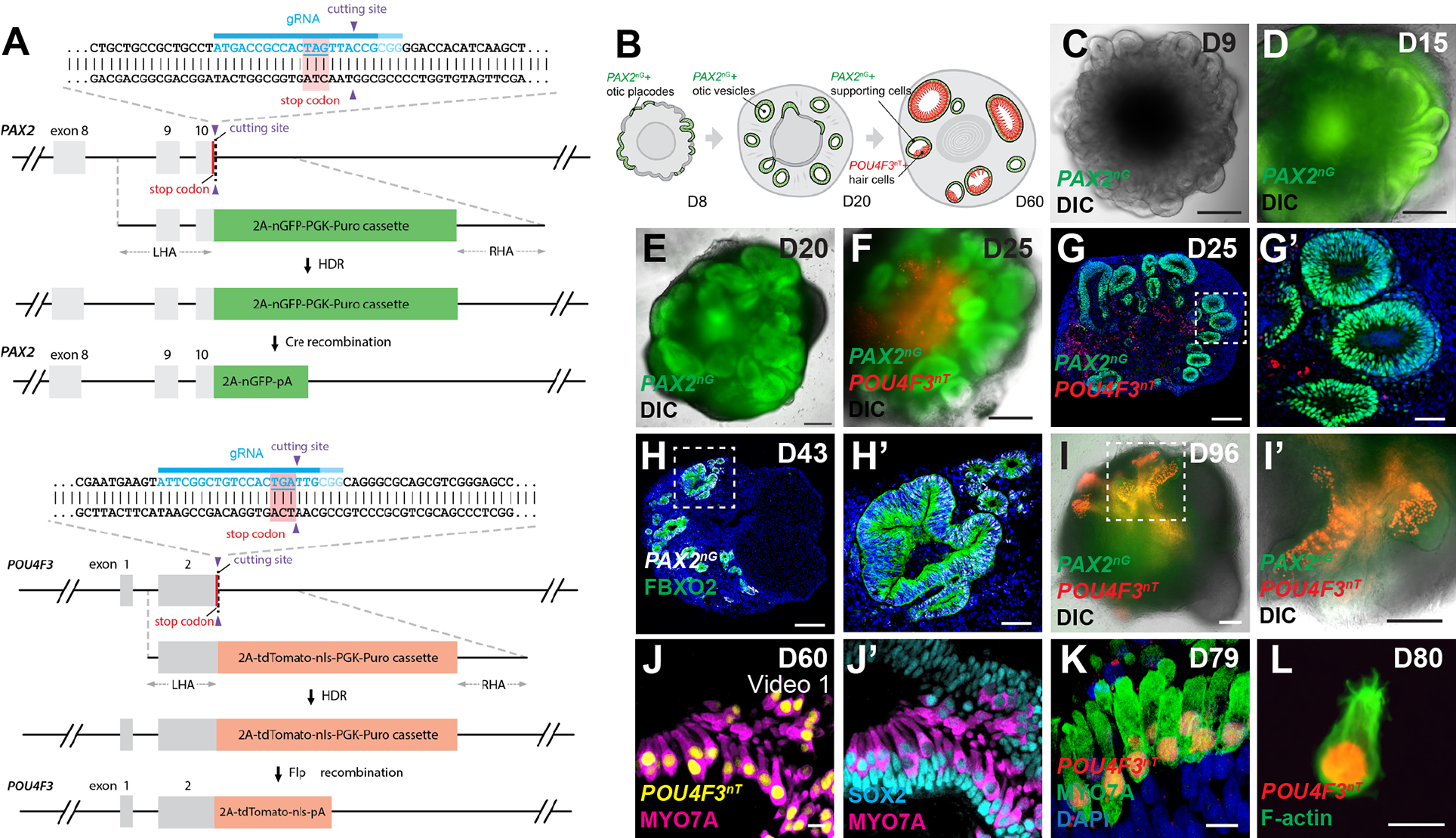
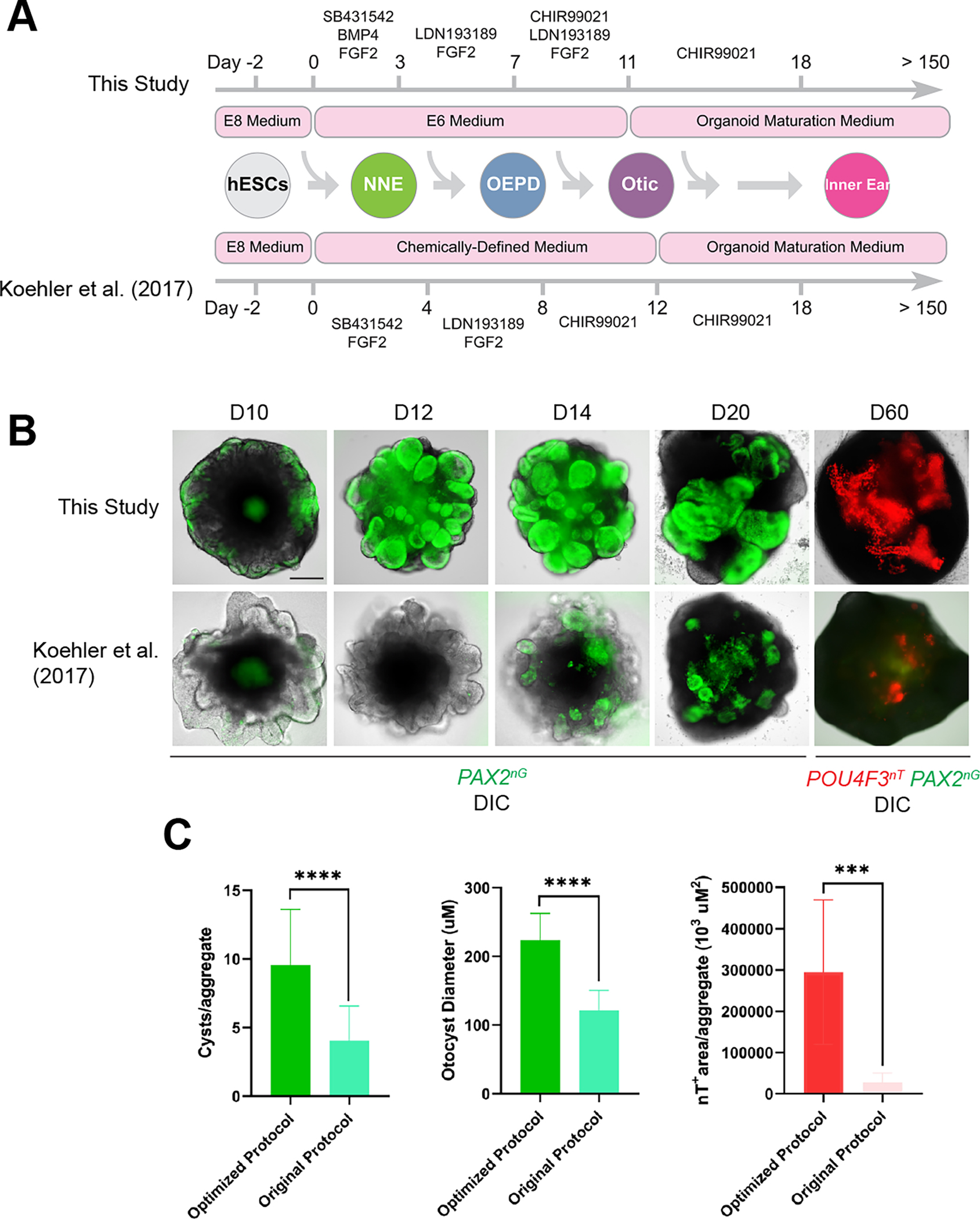
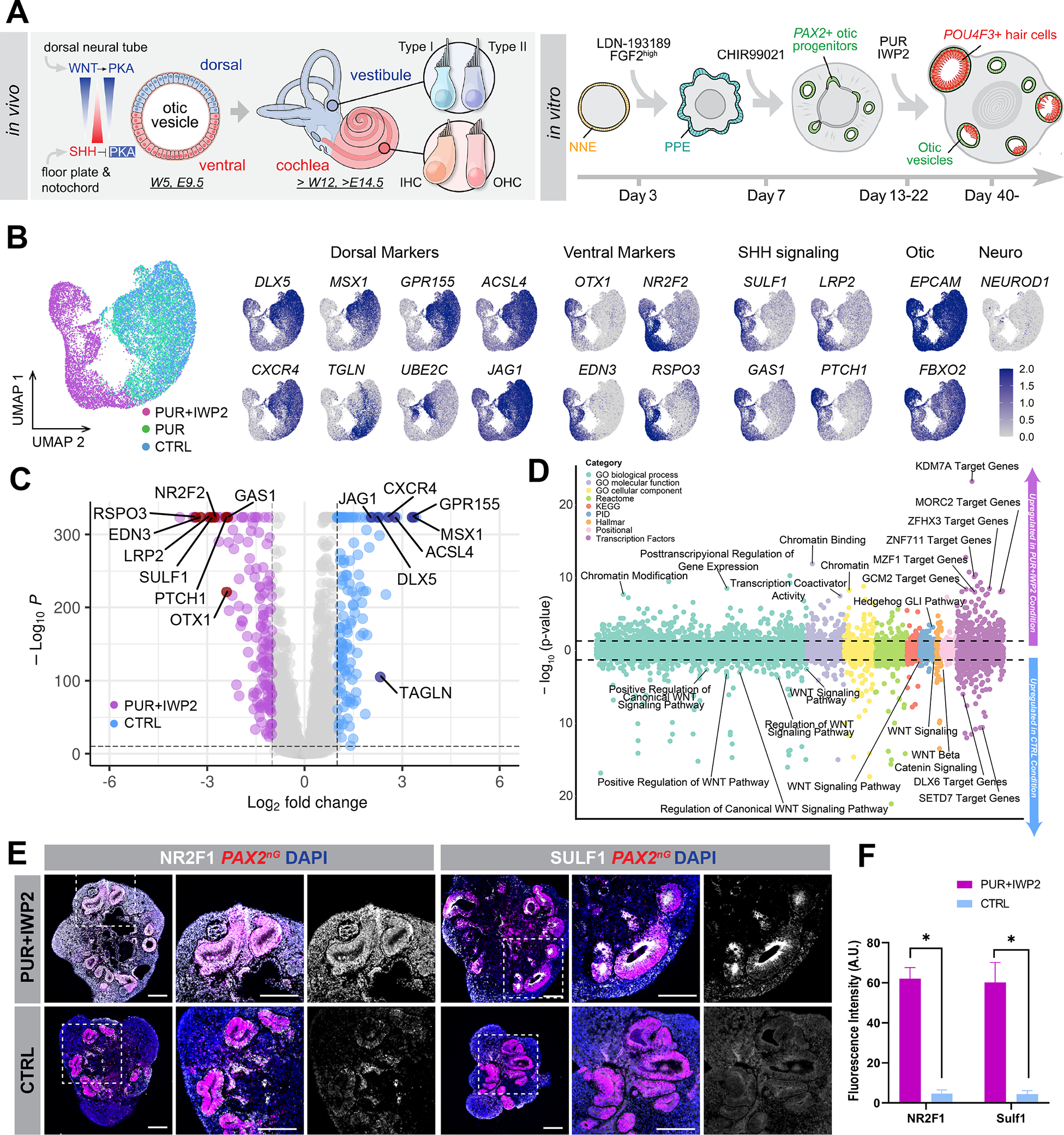
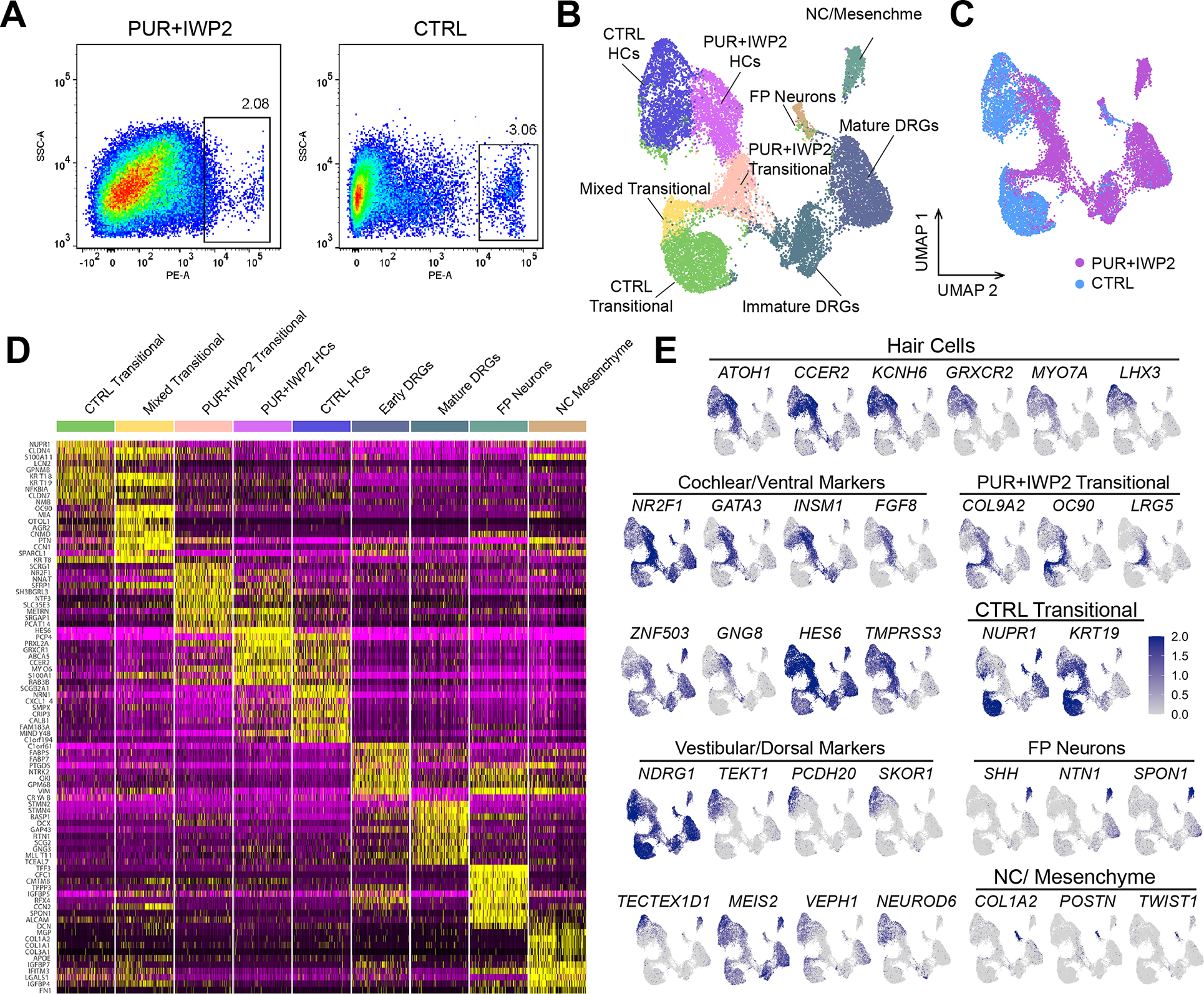
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
