

Organ function is preserved despite reorganization of niche architecture in the hair follicle
- PMID: 37419106
- PMCID: PMC10362479
- DOI: 10.1016/j.stem.2023.06.003
Organ function is preserved despite reorganization of niche architecture in the hair follicle
Abstract
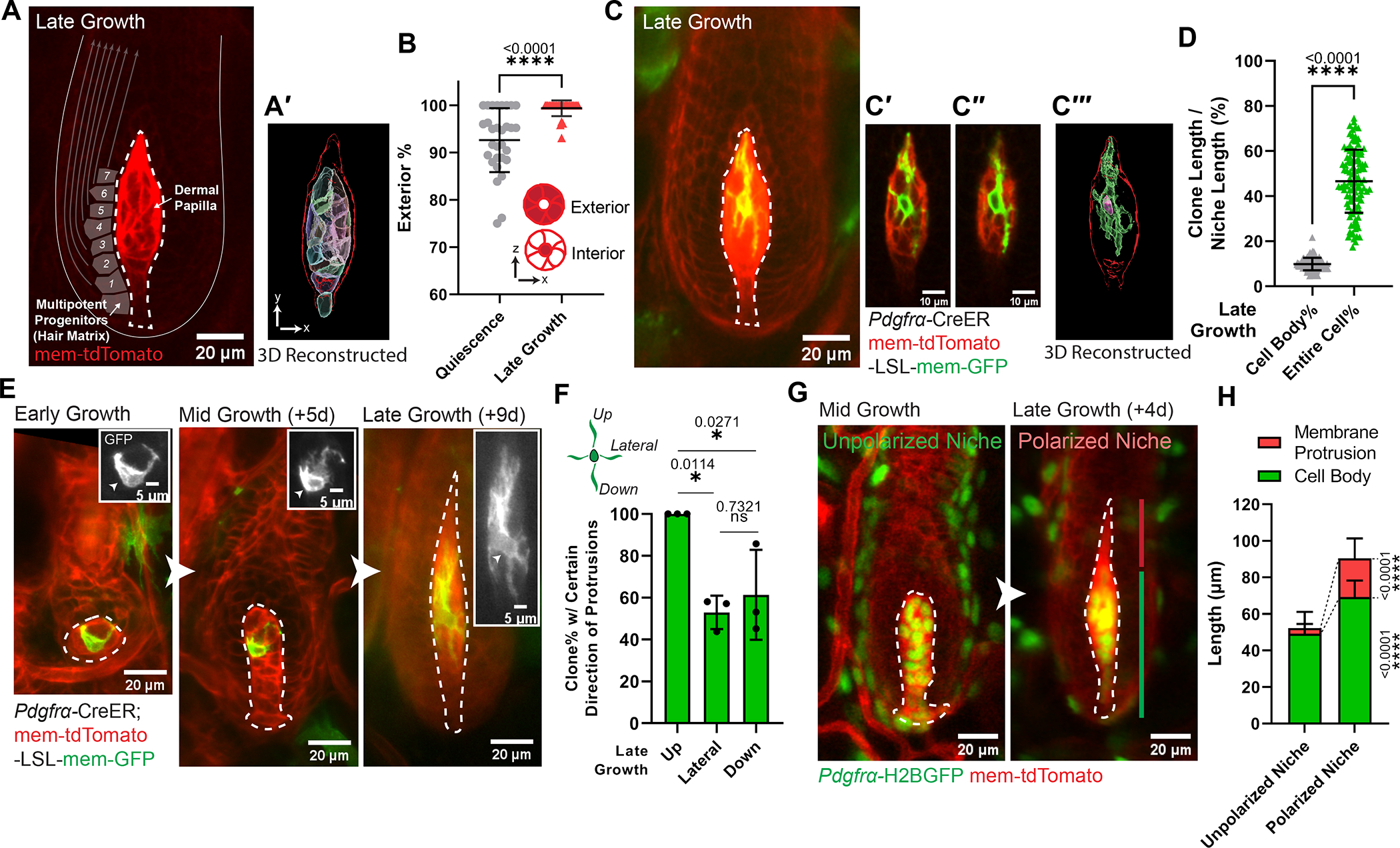
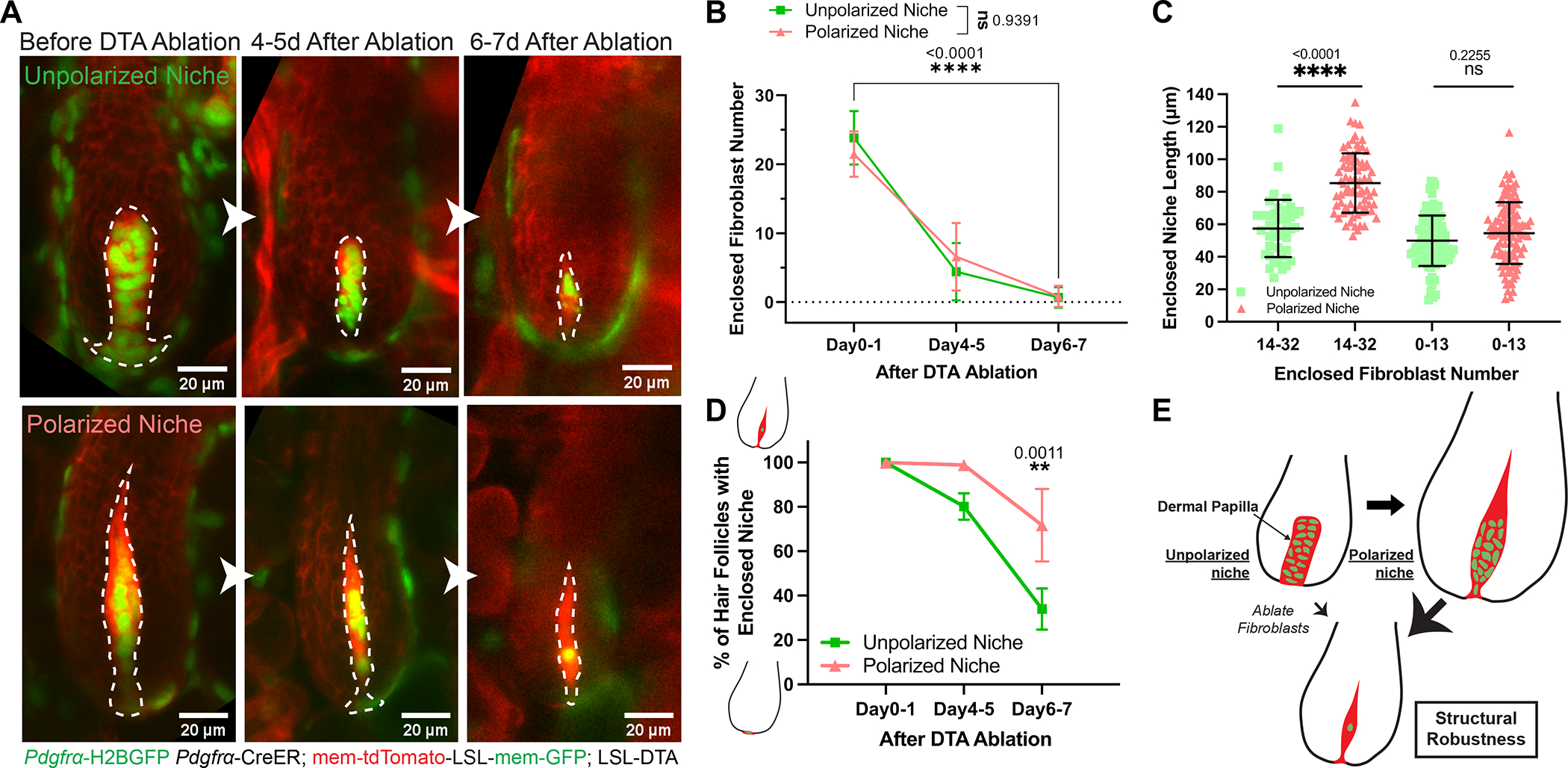
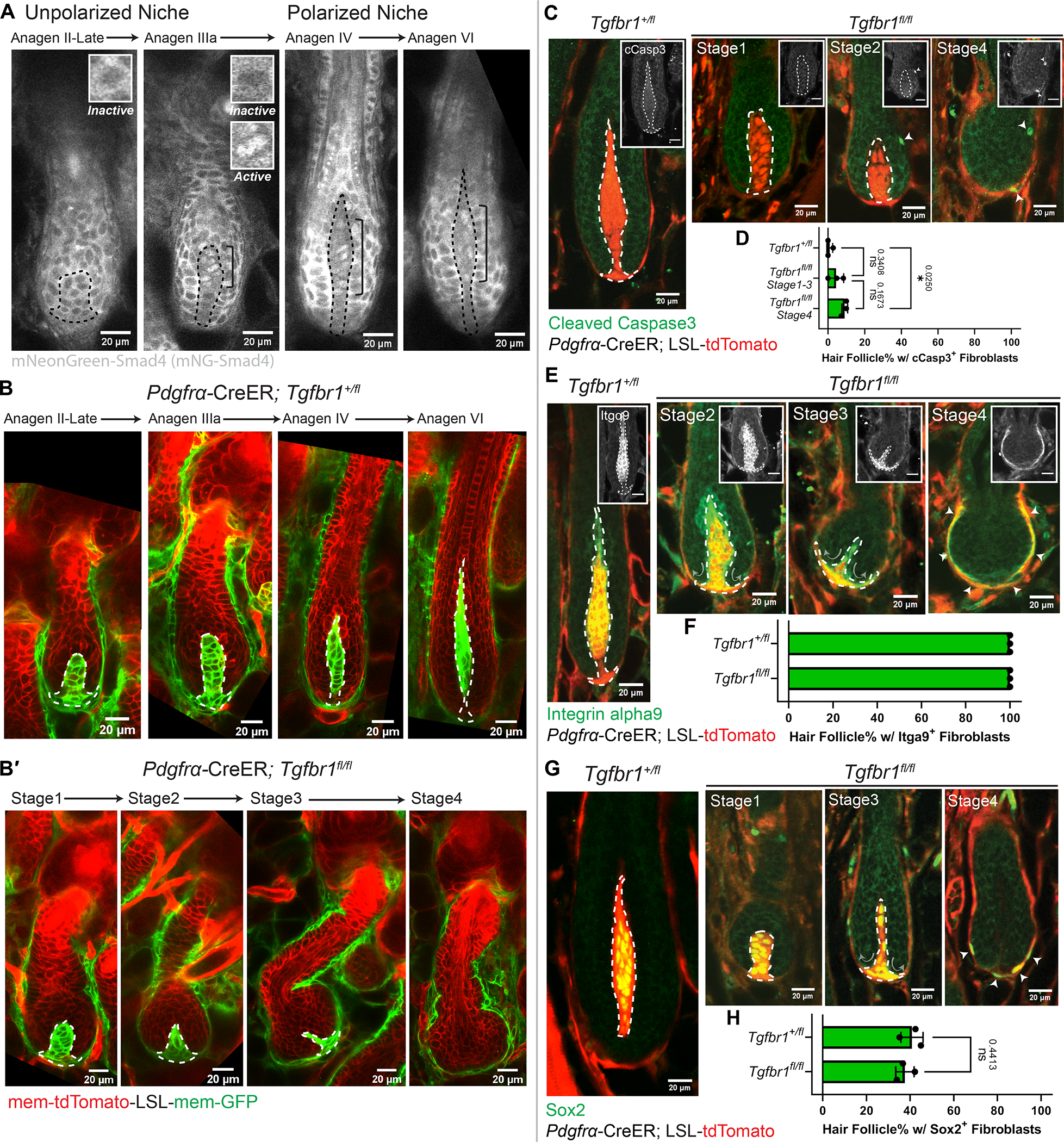
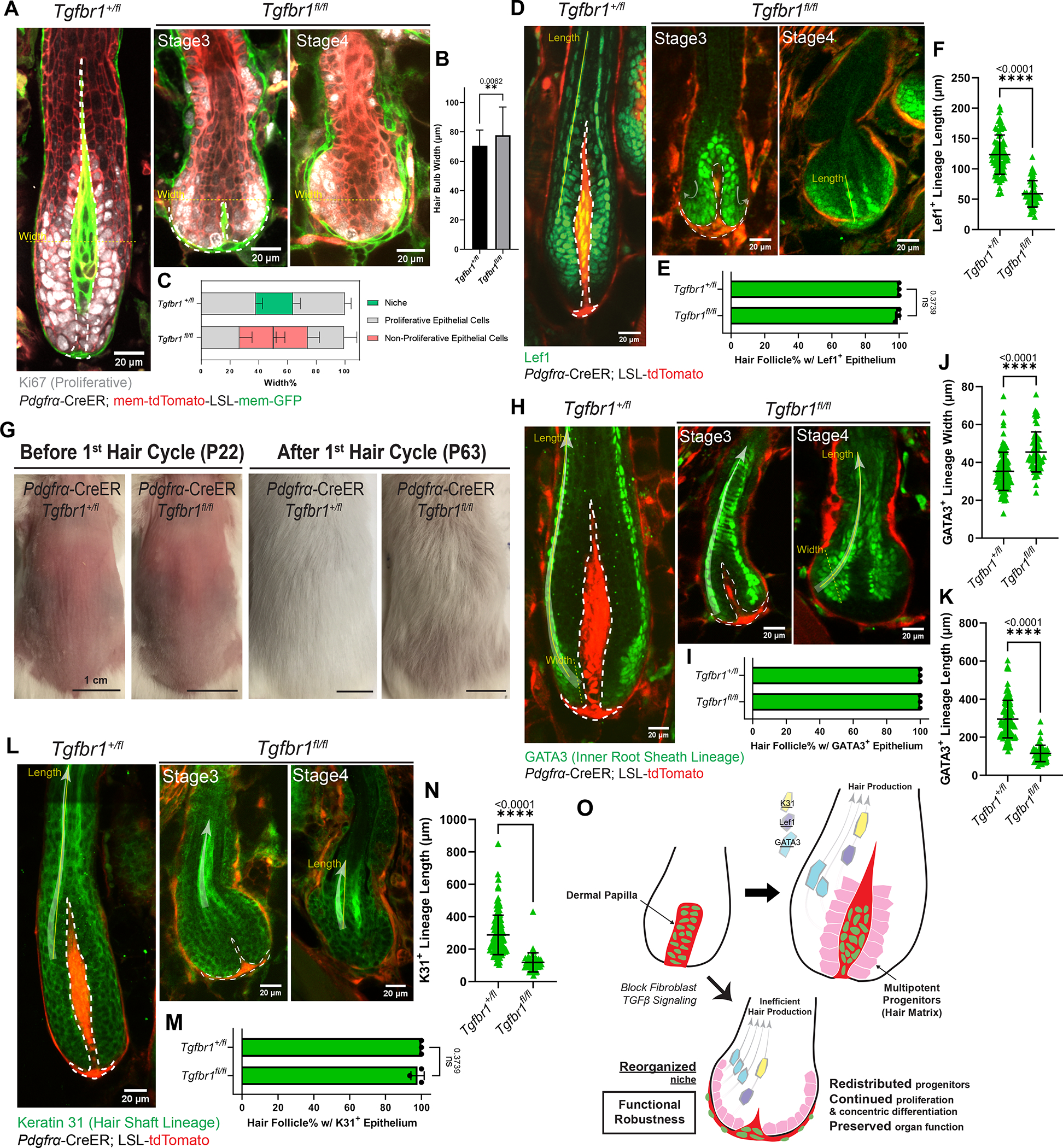
The ability of stem cells to build and replenish tissues depends on support from their niche. Although niche architecture varies across organs, its functional importance is unclear. During hair follicle growth, multipotent epithelial progenitors build hair via crosstalk with their remodeling fibroblast niche, the dermal papilla, providing a powerful model to functionally interrogate niche architecture. Through mouse intravital imaging, we show that dermal papilla fibroblasts remodel individually and collectively to form a morphologically polarized, structurally robust niche. Asymmetric TGF-β signaling precedes morphological niche polarity, and loss of TGF-β signaling in dermal papilla fibroblasts leads them to progressively lose their stereotypic architecture, instead surrounding the epithelium. The reorganized niche induces the redistribution of multipotent progenitors but nevertheless supports their proliferation and differentiation. However, the differentiated lineages and hairs produced by progenitors are shorter. Overall, our results reveal that niche architecture optimizes organ efficiency but is not absolutely essential for organ function.
Keywords: hair follicle; intravital imaging; regeneration; skin fibroblasts; stem cell niche; tissue architecture.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
