

Extreme summers impact cropland and grassland soil microbiomes
- PMID: 37419993
- PMCID: PMC10504347
- DOI: 10.1038/s41396-023-01470-5
Extreme summers impact cropland and grassland soil microbiomes
Abstract
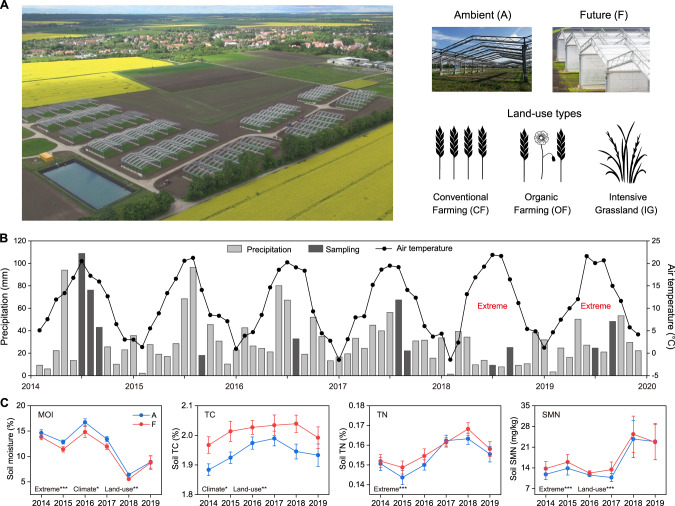
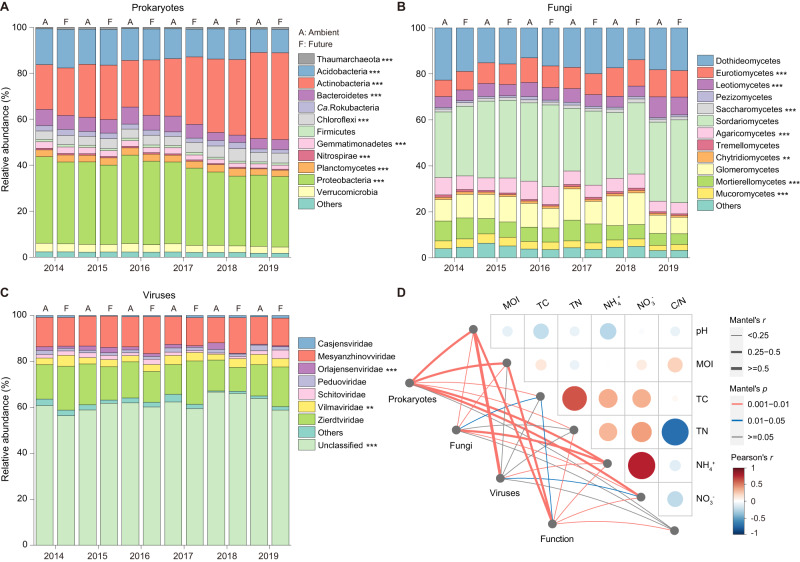
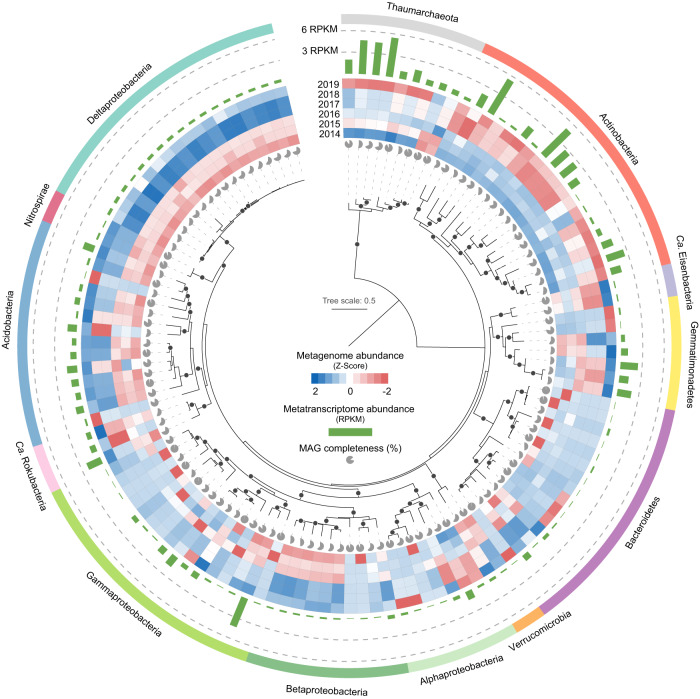
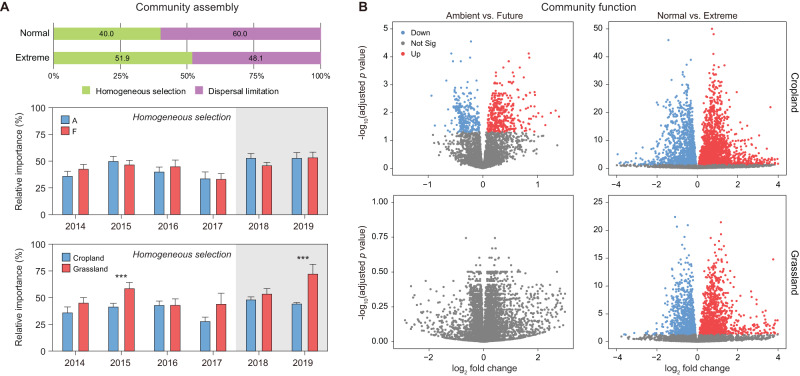
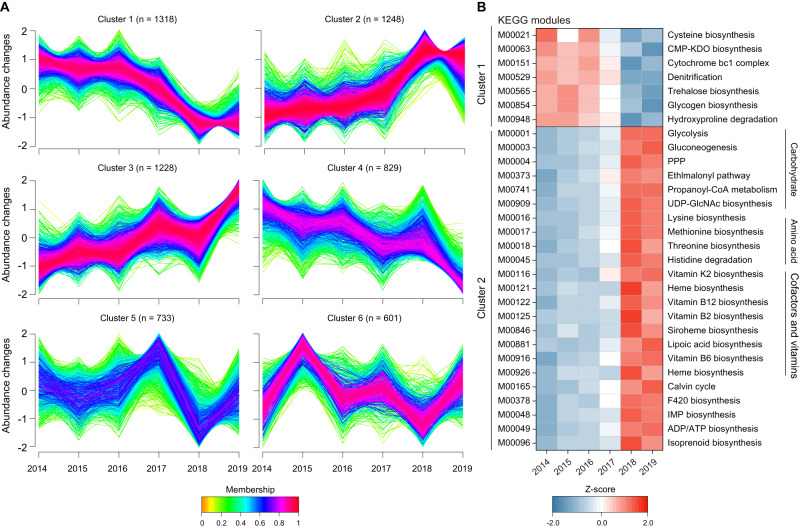
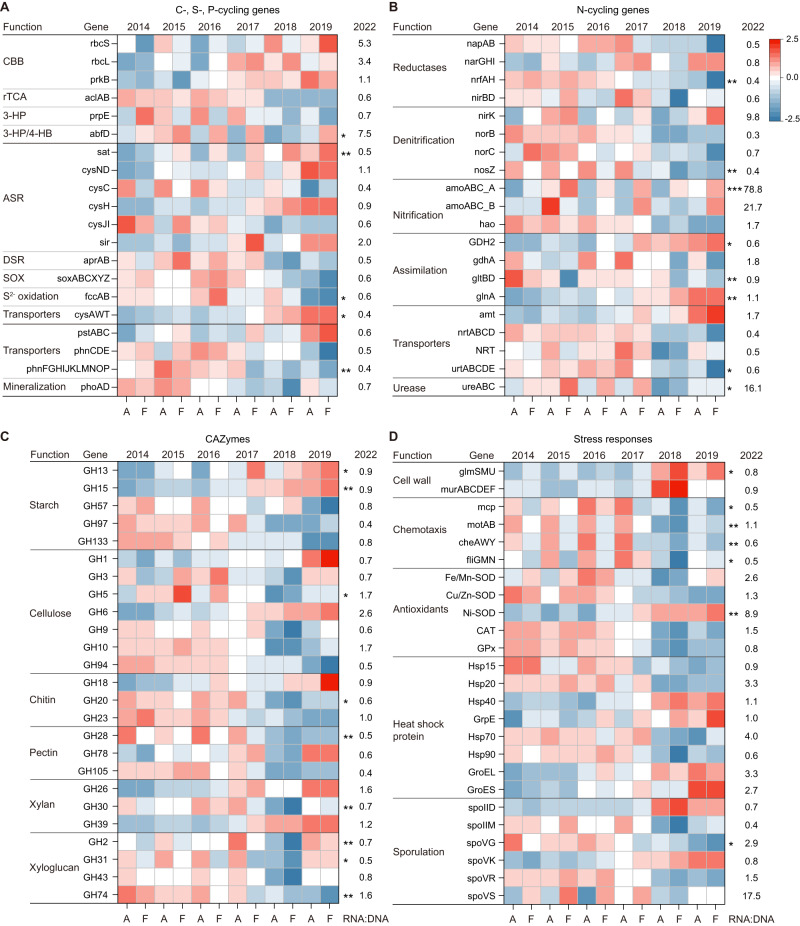
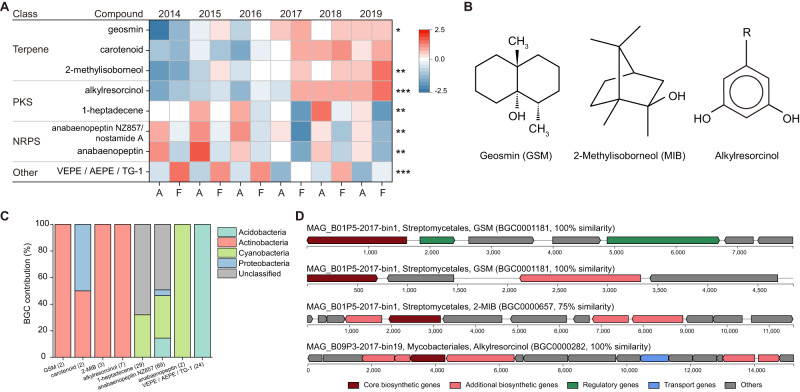
The increasing frequency of extreme weather events highlights the need to understand how soil microbiomes respond to such disturbances. Here, metagenomics was used to investigate the effects of future climate scenarios (+0.6 °C warming and altered precipitation) on soil microbiomes during the summers of 2014-2019. Unexpectedly, Central Europe experienced extreme heatwaves and droughts during 2018-2019, causing significant impacts on the structure, assembly, and function of soil microbiomes. Specifically, the relative abundance of Actinobacteria (bacteria), Eurotiales (fungi), and Vilmaviridae (viruses) was significantly increased in both cropland and grassland. The contribution of homogeneous selection to bacterial community assembly increased significantly from 40.0% in normal summers to 51.9% in extreme summers. Moreover, genes associated with microbial antioxidant (Ni-SOD), cell wall biosynthesis (glmSMU, murABCDEF), heat shock proteins (GroES/GroEL, Hsp40), and sporulation (spoIID, spoVK) were identified as potential contributors to drought-enriched taxa, and their expressions were confirmed by metatranscriptomics in 2022. The impact of extreme summers was further evident in the taxonomic profiles of 721 recovered metagenome-assembled genomes (MAGs). Annotation of contigs and MAGs suggested that Actinobacteria may have a competitive advantage in extreme summers due to the biosynthesis of geosmin and 2-methylisoborneol. Future climate scenarios caused a similar pattern of changes in microbial communities as extreme summers, but to a much lesser extent. Soil microbiomes in grassland showed greater resilience to climate change than those in cropland. Overall, this study provides a comprehensive framework for understanding the response of soil microbiomes to extreme summers.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Soil microbiomes show consistent and predictable responses to extreme events.Nature. 2024 Dec;636(8043):690-696. doi: 10.1038/s41586-024-08185-3. Epub 2024 Nov 27. Nature. 2024. PMID: 39604724 Free PMC article.
-
Sustainable Land Use Enhances Soil Microbial Respiration Responses to Experimental Heat Stress.Glob Chang Biol. 2025 Apr;31(4):e70214. doi: 10.1111/gcb.70214. Glob Chang Biol. 2025. PMID: 40272845 Free PMC article.
-
Drought consistently alters the composition of soil fungal and bacterial communities in grasslands from two continents.Glob Chang Biol. 2018 Jul;24(7):2818-2827. doi: 10.1111/gcb.14113. Epub 2018 Mar 26. Glob Chang Biol. 2018. PMID: 29505170
-
The Negative Legacy Effect of Extreme Drought on Soil Respiration Is Unaffected by Post-Drought Precipitation Regime in a Temperate Grassland.Glob Chang Biol. 2025 Feb;31(2):e70083. doi: 10.1111/gcb.70083. Glob Chang Biol. 2025. PMID: 39953799 Free PMC article.
-
Phyllosphere Community Assembly and Response to Drought Stress on Common Tropical and Temperate Forage Grasses.Appl Environ Microbiol. 2021 Aug 11;87(17):e0089521. doi: 10.1128/AEM.00895-21. Epub 2021 Aug 11. Appl Environ Microbiol. 2021. PMID: 34161142 Free PMC article.
Cited by
-
The influence of rhizosphere soil microorganisms and environmental factors on gentiopicroside content in the roots and rhizomes of Gentiana scabra Bunge from Liaoning Province.Front Microbiol. 2025 Mar 13;16:1554981. doi: 10.3389/fmicb.2025.1554981. eCollection 2025. Front Microbiol. 2025. PMID: 40182295 Free PMC article.
-
Climate extremes disrupt fungal-bacterial interactions.Nat Microbiol. 2023 Dec;8(12):2226-2229. doi: 10.1038/s41564-023-01531-7. Nat Microbiol. 2023. PMID: 38030901 No abstract available.
-
Impact of wine-grape continuous cropping on soil enzyme activity and the composition and function of the soil microbial community in arid areas.Front Microbiol. 2024 Feb 13;15:1348259. doi: 10.3389/fmicb.2024.1348259. eCollection 2024. Front Microbiol. 2024. PMID: 38414771 Free PMC article.
-
Increasing drying changes the relationship between biodiversity and ecosystem multifunctionality.NPJ Biofilms Microbiomes. 2025 May 6;11(1):72. doi: 10.1038/s41522-025-00711-z. NPJ Biofilms Microbiomes. 2025. PMID: 40328787 Free PMC article.
-
Harnessing co-evolutionary interactions between plants and Streptomyces to combat drought stress.Nat Plants. 2024 Aug;10(8):1159-1171. doi: 10.1038/s41477-024-01749-1. Epub 2024 Jul 24. Nat Plants. 2024. PMID: 39048724 Review.
References
-
- Fischer EM, Knutti R. Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes. Nat Clim Change. 2015;5:560–4. doi: 10.1038/nclimate2617. - DOI
-
- Schuldt B, Buras A, Arend M, Vitasse Y, Beierkuhnlein C, Damm A, et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl Ecol. 2020;45:86–103. doi: 10.1016/j.baae.2020.04.003. - DOI
-
- Toreti A, Belward A, Perez‐Dominguez I, Naumann G, Luterbacher J, Cronie O, et al. The exceptional 2018 European water seesaw calls for action on adaptation. Earth’s Future. 2019;7:652–63. doi: 10.1029/2019EF001170. - DOI
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Research Materials
