

ADAR1 Zα domain P195A mutation activates the MDA5-dependent RNA-sensing signaling pathway in brain without decreasing overall RNA editing
- PMID: 37421629
- PMCID: PMC10691306
- DOI: 10.1016/j.celrep.2023.112733
ADAR1 Zα domain P195A mutation activates the MDA5-dependent RNA-sensing signaling pathway in brain without decreasing overall RNA editing
Abstract
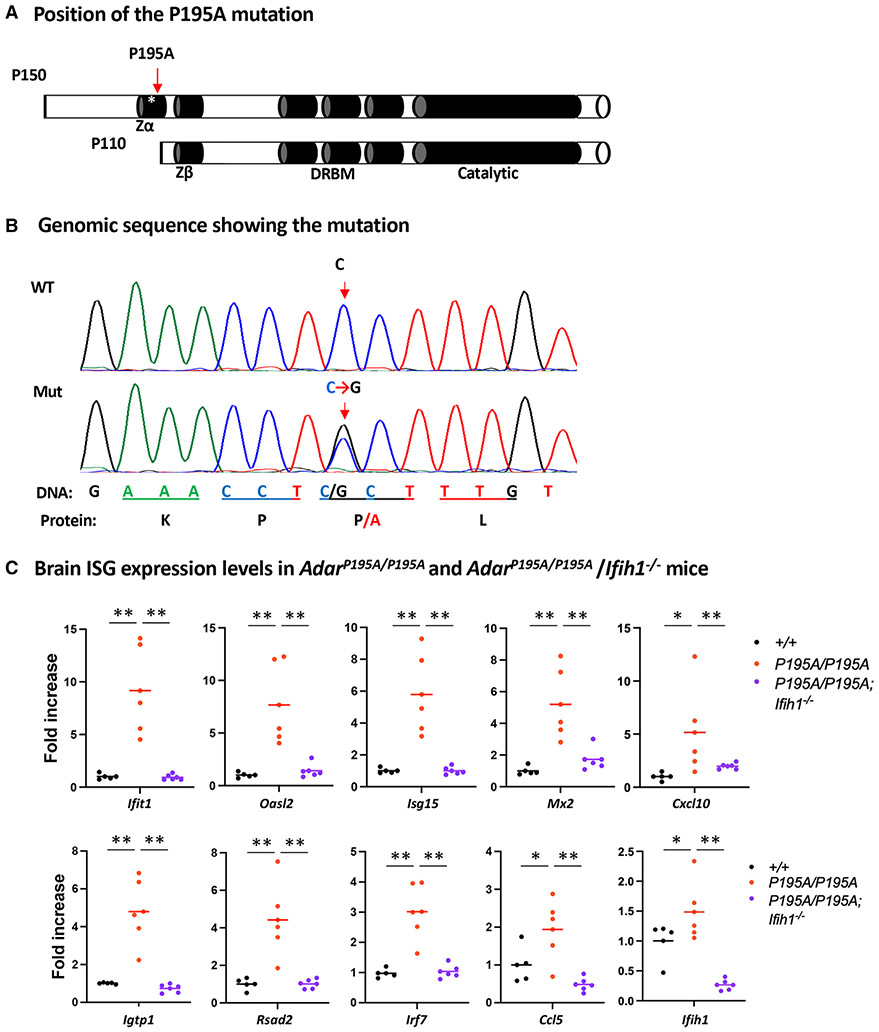
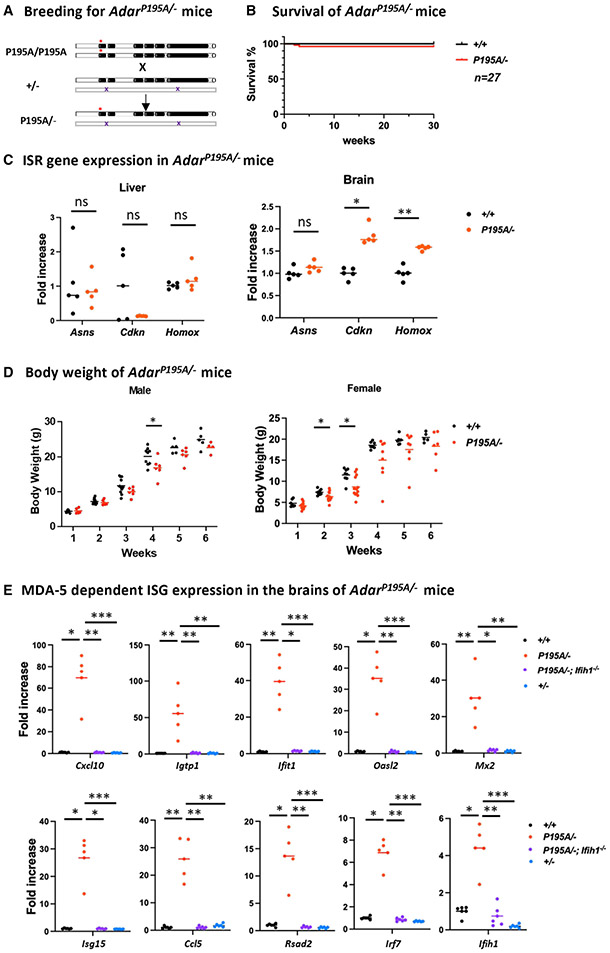
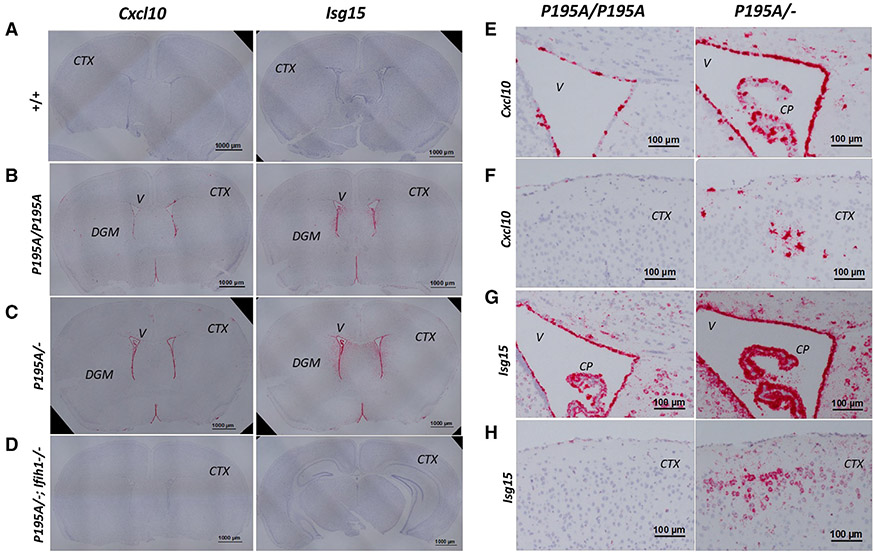
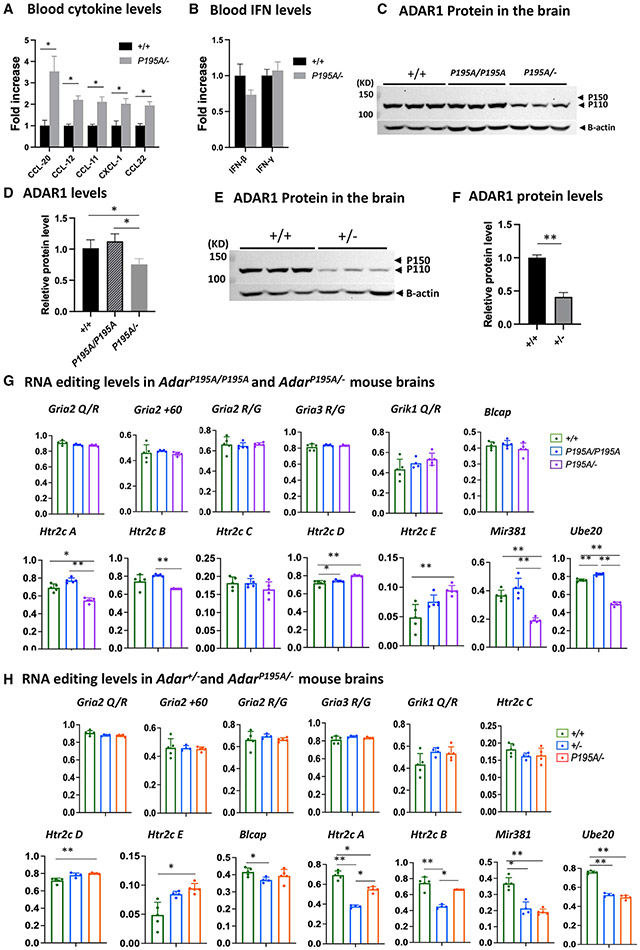
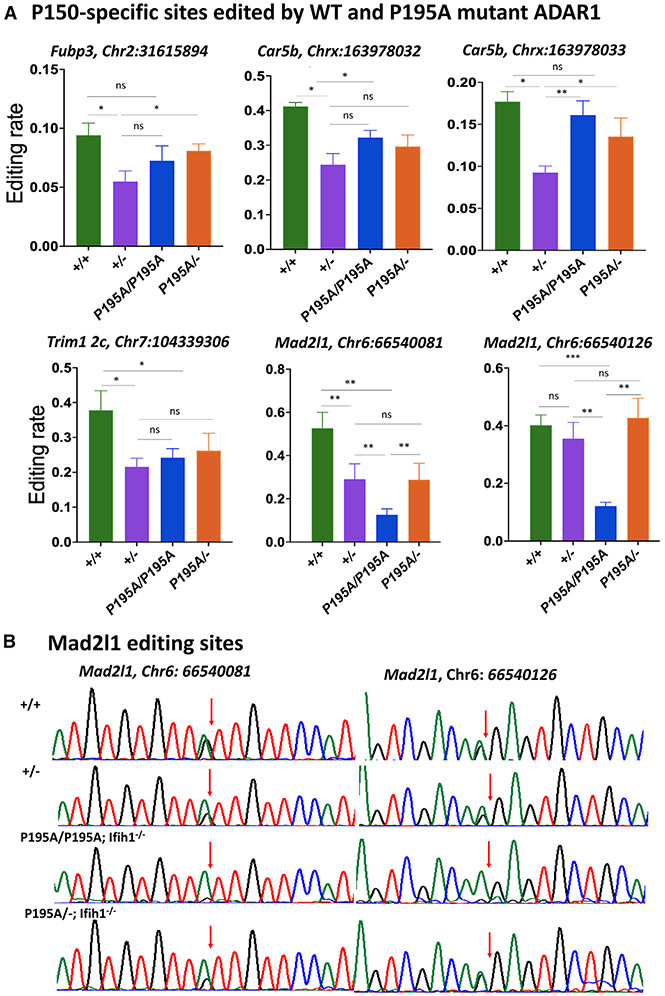
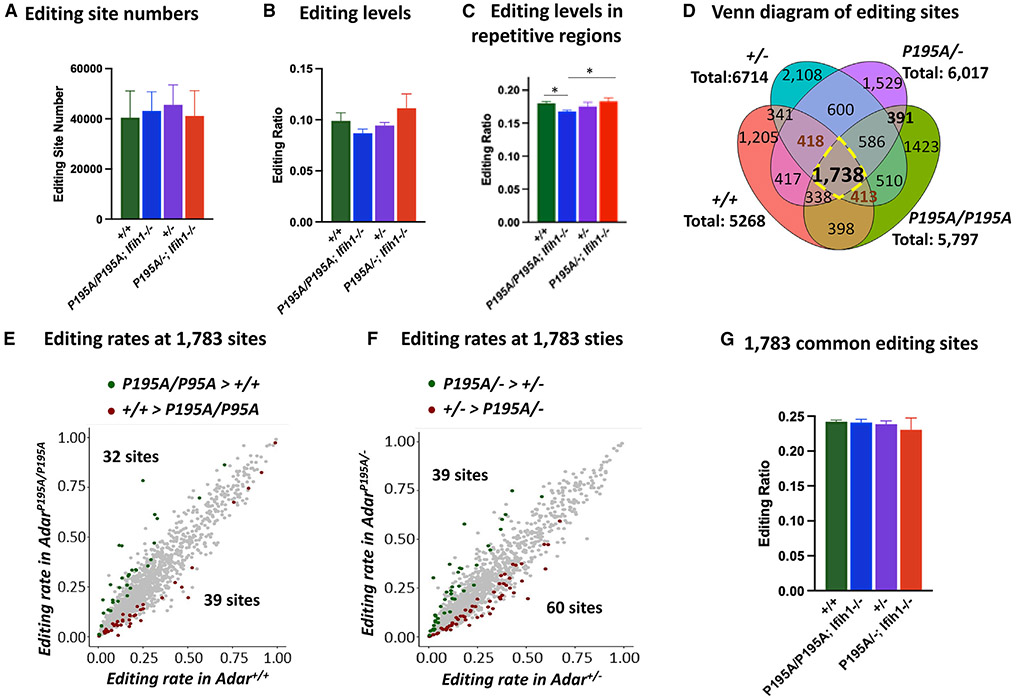
Variants of the RNA-editing enzyme ADAR1 cause Aicardi-Goutières syndrome (AGS), in which severe inflammation occurs in the brain due to innate immune activation. Here, we analyze the RNA-editing status and innate immune activation in an AGS mouse model that carries the Adar P195A mutation in the N terminus of the ADAR1 p150 isoform, the equivalent of the P193A human Zα variant causal for disease. This mutation alone can cause interferon-stimulated gene (ISG) expression in the brain, especially in the periventricular areas, reflecting the pathologic feature of AGS. However, in these mice, ISG expression does not correlate with an overall decrease in RNA editing. Rather, the enhanced ISG expression in the brain due to the P195A mutant is dose dependent. Our findings indicate that ADAR1 can regulate innate immune responses through Z-RNA binding without changing overall RNA editing.
Keywords: A-to-I RNA editing; ADAR1; AGS; Aicardi-Goutières syndrome; CP: Immunology; IFN-stimulated gene; ISG; MDA-5; RNA-sensing signaling pathway; Z-RNA; adenosine deaminase acting on RNA 1; melanoma differentiation-associated protein 5; neuroinflammation.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
The phenotype of the most common human ADAR1p150 Zα mutation P193A in mice is partially penetrant.EMBO Rep. 2023 May 4;24(5):e55835. doi: 10.15252/embr.202255835. Epub 2023 Mar 28. EMBO Rep. 2023. PMID: 36975179 Free PMC article.
-
Aicardi-Goutières syndrome-associated mutation at ADAR1 gene locus activates innate immune response in mouse brain.J Neuroinflammation. 2021 Jul 31;18(1):169. doi: 10.1186/s12974-021-02217-9. J Neuroinflammation. 2021. PMID: 34332594 Free PMC article.
-
The RNA-editing enzyme ADAR1: a regulatory hub that tunes multiple dsRNA-sensing pathways.Int Immunol. 2023 Mar 14;35(3):123-133. doi: 10.1093/intimm/dxac056. Int Immunol. 2023. PMID: 36469491
-
Deciphering the Biological Significance of ADAR1-Z-RNA Interactions.Int J Mol Sci. 2021 Oct 23;22(21):11435. doi: 10.3390/ijms222111435. Int J Mol Sci. 2021. PMID: 34768866 Free PMC article. Review.
-
ADAR RNA editing in innate immune response phasing, in circadian clocks and in sleep.Biochim Biophys Acta Gene Regul Mech. 2019 Mar;1862(3):356-369. doi: 10.1016/j.bbagrm.2018.10.011. Epub 2018 Oct 31. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 30391332 Review.
Cited by
-
Novel insights into double-stranded RNA-mediated immunopathology.Nat Rev Immunol. 2024 Apr;24(4):235-249. doi: 10.1038/s41577-023-00940-3. Epub 2023 Sep 26. Nat Rev Immunol. 2024. PMID: 37752355 Review.
-
A Wonderful Journey: The Diverse Roles of Adenosine Deaminase Action on RNA 1 (ADAR1) in Central Nervous System Diseases.CNS Neurosci Ther. 2025 Jan;31(1):e70208. doi: 10.1111/cns.70208. CNS Neurosci Ther. 2025. PMID: 39753993 Free PMC article. Review.
-
Neurodegenerative diseases reflect the reciprocal roles played by retroelements in regulating memory and immunity.Front Neurosci. 2024 Sep 20;18:1445540. doi: 10.3389/fnins.2024.1445540. eCollection 2024. Front Neurosci. 2024. PMID: 39371608 Free PMC article.
-
Deciphering the mechanistic roles of ADARs in cancer pathogenesis, tumor immune evasion, and drug resistance.Front Immunol. 2025 Aug 7;16:1621585. doi: 10.3389/fimmu.2025.1621585. eCollection 2025. Front Immunol. 2025. PMID: 40852728 Free PMC article. Review.
-
The Flipons, Infections, and Amyloids that Foreshadow the Fading Memories of Alzheimer's Disease.Neurosci Insights. 2025 Jun 6;20:26331055251338815. doi: 10.1177/26331055251338815. eCollection 2025. Neurosci Insights. 2025. PMID: 40486248 Free PMC article. Review.
References
-
- Yang S, Deng P, Zhu Z, Zhu J, Wang G, Zhang L, Chen AF, Wang T, Sarkar SN, Billiar TR, and Wang Q (2014). Adenosine deaminase acting on RNA 1 limits RIG-I RNA detection and suppresses IFN production responding to viral and endogenous RNAs. J. Immunol 193, 3436–3445. 10.4049/jimmunol.1401136. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
